
Variationen i det åbne land
Variation i rum: samfund og gradienter
Enhver, der færdes i naturen, vil have lagt mærke til, at visse plantearter ofte vokser side om side, således f.eks. blågrøn star, hjertegræs og vild hør. På samme måde optræder bestemte dyrearter sammen, f.eks. rørspurv og rørsanger.
Plantesamfund
Sådanne hyppigt genkommende artskombinationer har ført til den opfattelse, at der eksisterer egentlige plante- og dyresamfund med en artssammensætning, som er entydig og adskiller sig klart fra andre samfund, og som findes i naturen under bestemte økologiske forhold. For vegetationen er der udviklet en detaljeret systematik til at beskrive sådanne samfund svarende til den systematik, man benytter til at karakterisere plantefamilier, -slægter og -arter.
Umiddelbart synes en sådan inddeling af plantedækket i velafgrænsede samfund at kunne finde anvendelse i det åbne land, som i Danmark overvejende fremstår som velafgrænsede felter, mest i form af dyrkede marker, men også fragmenter af mose, eng, græsland og hede.
I et dyrket markfelt er plante- og dyrelivet som regel ret ensartet. Det skyldes, at man for at maksimere høstudbyttet gør forholdene så ensartede som muligt. Midler hertil er dræning, gødskning, mergling og anvendelse af pesticider. Helt ensartet er vegetationen og dyrelivet i det enkelte felt alligevel sjældent. Derom vidner de tætte bestande af valmuer, som ofte ses i udkanten af en kornmark.
Økologiske gradienter
I de felter i det åbne land, som ikke dyrkes, eller som kun udnyttes ekstensivt, er variationen i vækstbetingelserne som regel større. I de fleste tilfælde er der tale om en jævn variation. F.eks. aftager j ordens indhold af organisk stof og vand jævnt langs en linje vinkelret på en søbred med afstanden til søen. En sådan gradvis ændring af de økologiske forhold i én retning kaldes en økologisk gradient. Gradienten kan være svag eller stejl, nogle steder så stejl, at den har karakter af et „spring“.
Ser man bort fra sådanne spring og undersøger organismernes fordeling langs en jævn, økologisk gradient, vil man konstatere, at også artssammensætningen ændrer sig gradvist langs den samme linje. I nogle tilfælde domineres dele af en jævn økologisk gradient af en enkelt eller nogle få arter. Det kan derfor umiddelbart se ud, som om der er tale om velafgrænsede zoner. Dette kunne støtte teorien om eksistensen af egentlige samfund.
Tager man imidlertid alle arter i betragtning, og ikke kun de dominerende arter, viser det sig, at ændringerne i artssammensætningen langs gradienten sker gradvist (boks 3-5), og der er således ikke tale om velafgrænsede samfund med hver deres specifikke kombination af arter. Når man i det åbne land igen og igen støder på de samme kombinationer af arter, skyldes det ikke, at de har en fast tilknytning til hinanden. Det skyldes derimod, at de optræder sammen, fordi vækstbetingelserne de pågældende steder tilfredsstiller dem alle.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Tabellen viser arternes dækningsgrad (i %) i 30 kvadrater på hver 1 m2, som blev udlagt i forlængelse af hinanden langs en ret linje vinkelret på bredden af et stærkt eutrofieret vandhul på Nekselø – fra den åbne vandflade og indad. Der er kun medtaget arter, der forekom i mere end ét kvadrat, men i nederste række er det samlede antal arter anført. Bemærk, at arterne stort set forekommer uafhængigt af hinanden. Som det fremgår, synes vegetationen at være opdelt i velafgrænsede zoner af nogle få, dominerende arter (fra vandhullet og indefter drejer det sig om: smalbladet dunhammer, rank vinterkarse og almindelig rajgræs). De fleste af de øvrige, mindre dominerende arter vokser kun i en del af en „zone“, således findes stor nælde kun i en del af „rank vinterkarse-zonen“. Fløjlsgræs vokser både i „rank vinterkarse-zonen“ og i „almindelig rajgræs-zonen“. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Beskrivelse af vegetationen
I de tilfælde, hvor man opererer med samfund, er det som regel, fordi en inddeling i mindre enheder på grundlag af vegetationen – eller dyrelivet – kan være et praktisk redskab til at få overblik over naturens mangfoldighed.
En inddeling af vegetationen kan foretages alene på grundlag af forekomsten hhv. fraværet af bestemte arter. I mange tilfælde kan det imidlertid være nyttigt også at have et mål for den vægt, hvormed de enkelte arter indgår i vegetationen. Til en sådan, såkaldt kvantitativ, beskrivelse af vegetationen er der udviklet forskellige metoder, hvoraf bestemmelse af frekvens og dækningsgrad er de almindeligst anvendte (boks 3-6).
| To meget anvendte mål for enkeltarters betydning i vegetationen er frekvens (hyppighed) og dækningsgrad. Begge metoder forudsætter, at vegetationen er ensartet. Ved en frekvensbestemmelse udlægges et antal prøveflader, der som regel (her i Danmark) er cirkler med hver et areal på 0,1 m2 – en metode, der blev introduceret af botanikeren C. Raunkiær. For hver cirkel noteres de arter, der er rodfæstede i den. Frekvensprocenten (F-%) beregnes som det antal prøveflader, hvor arten noteres, i procent af samtlige prøveflader. Ved en dækningsgradsbestemmelse udlægges en til flere prøveflader, som ofte er noget større end de flader, der benyttes ved en frekvensbestemmelse. For hver art bedømmes det, hvor stor en del af prøvefladen den dækker, idet man forestiller sig, at planterne vokser så tæt som muligt. Dækningsgraden kan udtrykkes i procent, eller man kan anvende en anden skala, f.eks. Hult-Sernanders skala (semilogaritmisk), hvor forskellige dækningsintervaller tildeles en talværdi: 5: dækker mere end halvdelen af prøvefladen, 4: dækker mellem en fjerdedel og halvdelen af prøvefladen, 3: dækker mellem en ottendedel og en fjerdedel af prøvefladen, 2: dækker mellem en sekstendedel og en ottendedel af prøvefladen, 1: dækker mellem en toogtredivtedel og en sekstendedel af prøvefladen, + : dækker mindre end en toogtredivtedel af prøvefladen.
Begge metoder har fordele og ulemper. Frekvens-bestemmelsen er generelt mindre subjektiv end dækningsgrads-bestemmelsen, fordi det er forholdsvis entydigt at fastslå, om en art er rodfæstet inden for prøvefladen eller ej. Ved dækningsgrads-bestemmelsen kan der være meget stor forskel på forskellige personers vurdering. Dertil kommer, at arternes frekvens varierer meget mindre end deres dækningsgrad – både i løbet af en vækstsæson og gennem årene (men måske er det netop denne variation, man ønsker at beskrive). På den anden side giver frekvensbestemmelsen ofte et fortegnet billede af arternes relative betydning i vegetationen, idet små arter overvurderes og store arter undervurderes. Tabellen viser resultatet af en beskrivelse af overdrevsvegetationen på Glænø ved hjælp af både frekvens og dækningsgrad. En mere objektiv metode til bestemmelse af arternes dækningsgrad er pinpoint-analysen. På en kvadratisk ramme udspændes et antal snore (f.eks. 10 på den ene led og 10 på den anden led) med lige stor indbyrdes afstand, således at de danner et kvadratnet. Rammen placeres over vegetationen. En tynd metalstang føres lodret ned mod vegetationen i hvert af skæringspunkterne mellem de to sæt snore (i dette tilfælde 100 punkter) og det noteres, hvilke arter den rammer. Den enkelte arts dækningsgrad angives i % og beregnes som 100 x antal gange pinden ramte arten / antal skæringspunkter. |
|---|
Variation i tid: dynamik
Hvis man følger vegetationen og dyrelivet i et område i det åbne land over tid, vil man konstatere, at det er under bestandig forandring – både med årstiden og fra år til år. Under danske forhold spiller f.eks. temperatur og nedbør om foråret en stor rolle for vegetationens efterfølgende udvikling, især i naturtyper på tør bund. For mange dyr er vintertemperaturerne og sommerens længde vigtige for bestandenes størrelse. I disse tilfælde er der imidlertid blot tale om svingninger omkring et gennemsnit. Men der kan også ske retningsbestemte ændringer, som ikke kan sættes i forbindelse med tilfældige udsving i vejret. sådanne retningsbestemte ændringer over tid kaldes successioner (se nærmere i de følgende afsnit).
Bortset fra nogle få specielle tilfælde har det åbne lands naturtyper i Danmark det til fælles, at de befinder sig i en ustabil tilstand. Medmindre de løbende eller lejlighedsvis udsættes for indgreb eller påvirkninger, vil de med tiden udvikles til skov. Allerede i 1800-tallet beskæftigede danske botanikere sig med vegetationens dynamik, men først i anden halvdel af 1900-tallet er den enorme betydning af successionsprocesserne i det åbne land blevet åbenbar for alle. I dén periode begyndte virkningerne af de omfattende omlægninger af landbruget efter anden verdenskrig at vise sig, idet dyrkningen af den mest uproduktive landbrugsjord blev opgivet eller ekstensiveret, samtidig med at udnyttelsen af den resterende dyrkede jord blev intensiveret.
De fleste undersøgelser af successionsforløb har taget udgangspunkt i vegetationen, men også andre organismegrupper udviser ændringer gennem en succession. Man skelner mellem primære og sekundære successioner.
Afsnittet fortsætter efter boksen.
Boks

Forsøgsarealets udvikling fra 1983, ét år efter dyrkningens ophør, til 1995. De små kort viser bølget bunkes ændrede frekvens på arealet i de samme år.
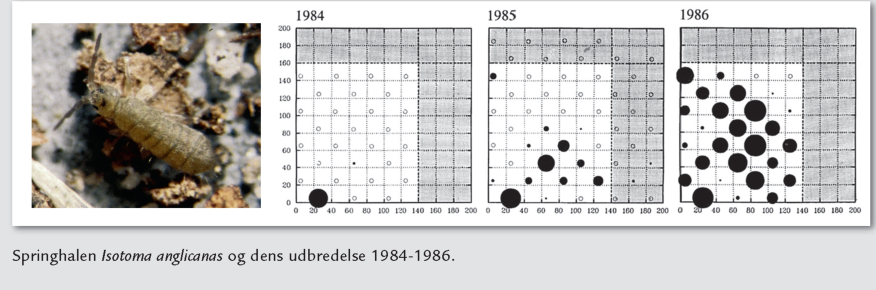
Springhalen Isotoma anglicanas og dens udbredelse 1984-1986.
| Forskere ved Naturhistorisk Museum og Aarhus Universitet iværksatte i 1979 en undersøgelse af en sekundær succession på Naturhistorisk Museums ejendom, Molslaboratoriet. Et ca. 2 ha stort sandet område med hedevegetation blev ryddet og dyrket med rug i to år. Et tilstødende areal blev udlagt som urørt kontrolområde. Forsøget efterlignede tidligere tiders græsmarksbrug, hvor kortvarig dyrkning af små marker blev fulgt af mange års braklægning. Efter høsten i efteråret 1982 blev området overladt til naturlig indvandring af planter og dyr fra den omgivende hede. Vegetationens sammensætning og udbredelse på arealet blev registreret før rydningen i sommeren 1979, i dyrkningsårene 1981-82 og derefter hver sommer fra 1983 til 1995. Foruden en registrering af arternes forekomst i 56 felter på arealet blev de almindelige arters frekvens og dækningsgrad (se ovenfor) i felterne målt. Der blev fundet 39 arter af karplanter før rydningen, og der var 40 i sommeren efter dyrkningens ophør. I årene derefter steg artsantallet og varierede fra 53 til 60 i perioden 1985-1993. I de sidste år af undersøgelsen, 1994-95, var der tendens til en nedgang. En- og toårige planter udgjorde en større procentdel af artsantallet i dyrkningsårene og de første år af successionen end senere i successionens forløb. Før rydningen var området helt domineret af græsarten bølget bunke og hedelyng samt opvækst af skov-fyr, eg og ene. Sommeren efter dyrkningens ophør var området næsten fuldstændig dækket af krybende hestegræs iblandet pletter med rødknæ. Disse planter prægede fortsat vegetationen i de følgende 2-3 år. Små frøplanter af hedelyng var allerede almindelige i anden sommer efter successionens start, men først nogle år senere var lyngens dækningsgrad så høj, at den begyndte at præge området. Den højeste dækningsgrad blev nået 9 år efter succes sionens start. Bølget bunke bredte sig langsomt fra kanten indefter i området (se kort), og så sent som 13 år efter successionens start var der kun få individer i områdets centrale del. Dyrelivets succession blev undersøgt ved hjælp afjordprøver og fælder beregnet på fangst af insekter og andre leddyr. Blandt jordbundens mikroleddyr var pansermiderne den gruppe, som var stærkest påvirket af dyrkningen og var længst om at genetablere deres bestande efter dyrk ningens ophør. Fedtmider og fløjlsmider var mindst påvirket, mens springhale- og rovmidebestanden relativt hurtigt nåede samme tæthed som før opdyrkningen. Der var dog store forskelle imellem springhalearterne. Nogle kom sig hurtigt, andre blev i de første successionsår mere almindelige end i kontrolområdet, og atter andre bredte sig meget langsomt ind i området fra omgivelserne. |
|---|
Afsnit fortsætter her.
Primære successioner
Primære successioner tager deres begyndelse på steder, hvor der ikke tidligere har været noget plantedække og heller ikke er nogen jordbund, altså ikke noget organisk stof. Det kan f.eks. være en sandstrand, en nøgen klippe eller en grusgrav. Udgangspunktet er altså det rå, geologiske materiale og de organismer, som spredes til stedet og etablerer sig. De første successionsstadier rummer alger, laver, mosser og enårige planter. Disse organismer opbygger en pulje af organisk stof. Dermed er vejen banet for dyr og mikroorganismer, der kan udnytte dette organiske stof. Bakterier, svampe og forskellige grupper af hvirvelløse dyr, f.eks. protozoer, rundorme, dværgregnorme, mider og springhaler, indvandrer meget hurtigt efter, at de første planter har etableret sig. Disse dyrs ekskrementer udgør en væsentlig del af den første jordbund, der dannes i begyndelsen af den primære succession (figur 3-20).
I de første successionsstadier afhænger plantedækkets sammensætning i høj grad af, hvilke arter der har mulighed for at spredes dertil, om de kan etablere sig, og om de i det hele taget kan klare sig i et miljø med stærkt svingende temperatur og fugtighed.
Efterhånden, som plantedækket sluttes, og en tættere vegetation af flerårige planter etablerer sig, modificeres mikroklimaet og jordbunden. Derved bliver miljøet bedre egnet som levested for plante- og dyrearter, som nok kunne sprede sig til området, men som ikke kunne klare sig under de barskere forhold, der herskede i de første successionsstadier. Endvidere begynder konkurrence at få betydning – i nogle tilfælde giver det sig udslag i, at allerede etablerede arter hæmmer etableringen af nye arter. På næringsrig bund kan en tæt, højtvoksende græs- eller højstaudevegetation således forsinke etableringen af buske og træer og dermed udviklingen mod skov.
I de senere stadier af tilvoksningen tager buske og lyskrævende træer over, og successionen slutter med skov, som er klimaksvegetationen under danske forhold. Et sådant successionsforløb tager nogle hundrede år. Samtidig med at vegetationen ændrer sig, sker der en jordbundsudvikling. Udviklingen fra sandstrand over hvid klit, grå klit og klithede til nåleskov er et eksempel på en primær succession.
Når der i vore dage er åbent land, hvor der ellers ville være skov, er årsagen som beskrevet nærmere i kapitlet Naturen i det åbne land (se Danmark er et potentielt skovland og de følgende afsnit) i langt de fleste tilfælde, at vegetationen fastholdes på et tidligt successionstrin. Det kan skyldes, at skoven – måske for mange hundrede år siden – er blevet fældet. Derved er successionen sat tilbage til et tidligere stadium. At det åbne land så ikke er sprunget i skov igen, kan som regel tilskrives græsning eller dyrkning. Hvis disse påvirkninger ophører, fortsætter successionen mod klimaksstadiet. Kun ganske få steder er naturens egne processer årsag til vedvarende skovløshed. Det er f.eks. tilfældet på stejle skrænter, på saltpåvirket bund og på fugtig bund med en stærkt svingende vandstand.
At successionen stopper som følge af påvirkninger fra menneskets side betyder imidlertid ikke, at der er tale om en fuldstændig stilstand. Eksempelvis er såvel et græsset overdrev som en dyrket mark dynamiske systemer. Der kan ske indvandring og etablering af nye arter, og andre arter uddør måske lokalt. Dertil kommer, at der – ud over de ovenfor omtalte svingninger – kan ske generelle, mere permanente ændringer i miljøet, f.eks. klimaændringer og luftforurening, alt sammen med ændringer i de pågældende naturtypers plante- og dyreliv til følge. Disse ændringer kan evt. ændre områdernes struktur og funktion så meget, at der fremkommer en helt anden naturtype.
Sekundære successioner
Sekundære successioner adskiller sig fra primære successioner ved, at de forløber på steder, hvor der forud har været et vegetationsdække og derfor allerede er udviklet en jordbund. Det kan f.eks. være marker, hvor dyrkningen er ophørt, således som det er tilfældet på sandede jorde mange steder i Danmark (boks 3-7). I jorden er der en pulje af organisk stof og næringsstoffer, og der kan ligge frø, rødder og jordstængler, så jorden hurtigt på ny dækkes af vegetation. Svampe, bakterier og jordbundsdyr er også til stede lige fra begyndelsen. Der er derfor mulighed for, at en sekundær succession i de indledende stadier forløber hurtigere end en primær succession, mens der i de senere stadier ikke er de store forskelle.
Successionsundersøgelser
Successionsforløb, som kan strække sig over århundreder, er i sagens natur vanskelige at undersøge. Principielt er gentagne undersøgelser af ét og samme område det eneste rigtige, når vegetationens dynamik skal beskrives. Men der foreligger fra det åbne land kun få undersøgelser, som strækker sig over meget lang tid – de bedste danske eksempler er Gammelmose syd for Lyngby (fra 1844), Nørholm Hede (fra 1921, se kapitlet Lynghederne: Præsentation og de følgende afsnit) og opgiven landbrugsjord på Fredmarken på øen Vorsø i Horsens Fjord (fra 1933).
En alternativ metode er på én og samme tid at undersøge arealer med vegetation af forskellig alder. Det kan f.eks. være marker, hvor dyrkningen er opgivet på forskellige tidspunkter. Her kan det imidlertid være et problem, hvis forholdene på de opgivne marker i udgangssituationen ikke har været helt ens, eller at der i den periode, der betragtes, er sket ændringer i det omgivende miljø, eksempelvis mht. klima, luftforurening eller indvandring af invasive arter.
Vejviser
Værket Naturen i Danmark i fem bind udkom i årene 2006-2013. Teksten ovenfor er kapitlet Variationen i det åbne land.
- Forrige afsnit er Symbiose
- Næste afsnit er Moserne og de ferske enge
Kommentarer
Kommentarer til artiklen bliver synlige for alle. Undlad at skrive følsomme oplysninger, for eksempel sundhedsoplysninger. Fagansvarlig eller redaktør svarer, når de kan.
Du skal være logget ind for at kommentere.