
Mange kan lide at gå på svampejagt i efterårsskoven. Andre fascineres med en vis skræk af det mylder af paddehatte, der efter kraftig sensommerregn pludselig varsler efterår. De iøjnefaldende paddehatte er dog kun et vidnesbyrd om svampenes liv, der langt overvejende foregår i det skjulte. Svampe er grundlæggende opbygget af mikroskopiske celletråde, der kaldes hyfer. Disse hyfer har en tykkelse på ca. 10 μm (1/100 mm), men danner vidt udstrakte netværk dér, hvor svampene lever, f.eks. i skovbunden, i råddent træ eller inde i levende planter. Et sådant hyfenetværk kaldes et mycelium og udgør den egentlige svamp (figur 6-1).
Svampene
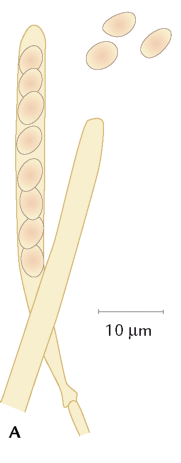
FIGUR 6-10 (a). To sporesække, den højre med afskudte sporer.
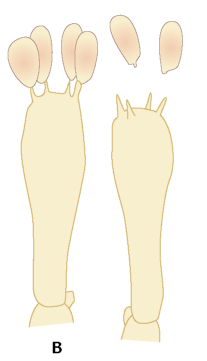
FIGUR 6-10 (b). To basidier med sporer; på den højre er de netop afskudt ved den mekanisme, som vises i detaljer nedenfor (figur 6-10 (c)).
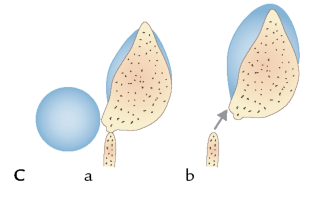
FIGUR 6-10 (c). Mekanisme for afskudte sporer.
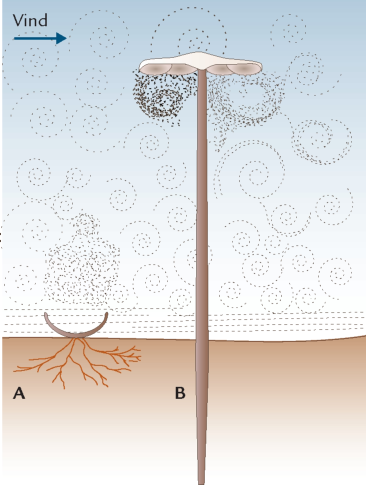
FIGUR 6-11. Forskellen i sporefrigørelse hos sæksvampe og basidiesvampe afspejles i frugtlegemernes udformning. Hos sæksvampene (A) vender sækkene opad, typisk i skålformede frugtlegemer eller indesluttet i flaskeformede frugtlegemer med opadvendt spids, således at sporerne kan skydes væk fra de stillestående luftlag nær skovbunden (vandret skravering) og op i de turbulente luftmasser (cirkulære markeringer). Hos basidiesvampene (B) vender basidierne derimod horisontalt eller med spidsen nedad, således at sporerne ikke bare falder ned på hymeniet igen efter frigivelsen. Samtidig har frugtlegemerne ofte en stilk, for at hymeniet kan komme op i en højde, hvor de frigjorte sporer falder ned i de turbulente luftlag, og en hat, der beskytter hymeniet mod nedbør, der ellers kan forstyrre sporefrigørelsen.

FIGUR 6-12 (a). Rør, lameller og pigge er forskellige måder at opnå den forøgelse af overfladen på, som ses hos henholdsvis rørhatte (A), lamelsvampe (B) eller pigsvampe (C). Der findes yderligere et utal af mærkværdige frugtlegemetyper hos basidiesvampene, der bryder med disse overordnede mønstre, f.eks. støvbolde, koralsvampe og bævresvampe.

FIGUR 6-12 (b). Lamelsvampe.

FIGUR 6-12 (c). Pigsvampe.

FIGUR 6-13. Brunstokket rørhat er en god spisesvamp og danner mykorrhiza med mange forskellige træarter. Den er almindelig i både løv- og nåleskov på morbund.

FIGUR 6-5 (a). Skæl-rørhat (Leccinum) er et eksempel på en svampeslægt med ektomykorrhiza og en høj grad af værtspecialisering. Rød birke-rørhat (A) findes således kun med birk, og orange aspe-rørhat (B) vokser udelukkende med bævreasp.

FIGUR 6-5 (b). Orange aspe-rørhat vokser udelukkende med bævreasp.

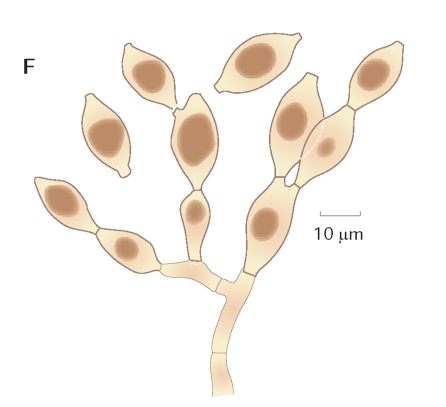
FIGUR 6-6. Spredning med ukønnede sporer ses hos mange forskellige svampe. Hos halvkugleformet kulbær dannes der store mængder af ukønnede sporer fra grå tottede, støvede strukturer (A), der siden gennemvokses af de rustrøde stroma, der indeholder kønnede frugtlegemer (B). Under mikroskopet ses de grenede hyfestrukturer, hvorfra de ellipseformede sporer frigives (C). Også hos barksvampen Botryobasidium aureum finder man ofte ukønnet og kønnet sporedannelse side om side. De kønnede sporer dannes i det tynde hvide spind (D), mens de gullige puder (E) er dannet af ukønnede citronformede sporer, der i massevis frigøres fra grenede hyfer (F).


FIGUR 6-7. Svampesporer kan være særdeles forskelligt udformet med en størrelse, der typisk varierer fra 5 til 50 μm. De fleste sporer er tilpasset spredning med luftstrømme, og deres ringe størrelse betyder, at de kan svæve i luften og blive spredt over meget lange afstande med vinden (A). Dette forhold kan formodentlig forklare, at mange svampe har en meget vid, i visse tilfælde global, udbredelse. Der findes dog også svampe, hvis sporer først og fremmest spredes gennem vand (B). Det er ikke alle sporetyper, der er beregnet til spredning. Mange svampe danner således tykvæggede hvilesporer, der først og fremmest sikrer, at svampen kan overleve lokalt i en periode med ugunstige forhold, f.eks. stærkt udtørring eller mangel på levende værtsplanter i vinterhalvåret (C).

FIGUR 6-8 (a). Almindelig stinksvamp (Phallus impudicus) danner sine sporer i en stærkt lugtende, kulhydratholdig og olivengrøn sporemasse, som tiltrækker mange forskellige insekter, ikke mindst fluer (A). Når fluerne æder af sporemassen, klæber nogle af sporerne til fluernes ben, hvorved de spredes til nye mulige voksesteder, når fluen flyver videre. En lignende spredningsstrategi anvendes i slægten Ophiostoma (B). Her udskilles både kønnede og ukønnede sporer i en slimet, kulhydratholdig væske, der ædes af eller klæber til forskellige insekter.

FIGUR 6-8 (b). I slægten Ophiostoma udskilles både kønnede og ukønnede sporer i en slimet, kulhydratholdig væske, der ædes af eller klæber til forskellige insekter.
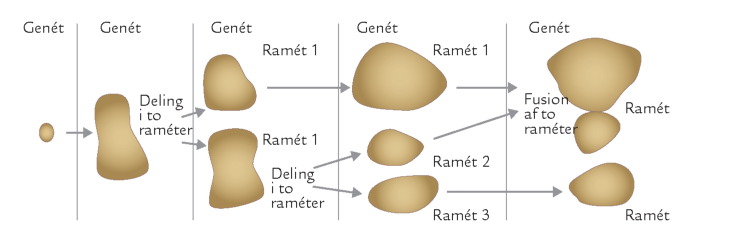
FIGUR 6-9. Et svampe-genét kan uden videre splittes op i flere selvstændigt fungerende, men genetisk ens mycelier, der kaldes raméter. Når to sådanne raméter mødes igen, kan de uden videre igen fusionere til et funktionelt mycelium. Hos visse svampe, der overvejende spredes ved ukønnede sporer, kan raméter inden for en enkelt genét være spredt over hele verden.
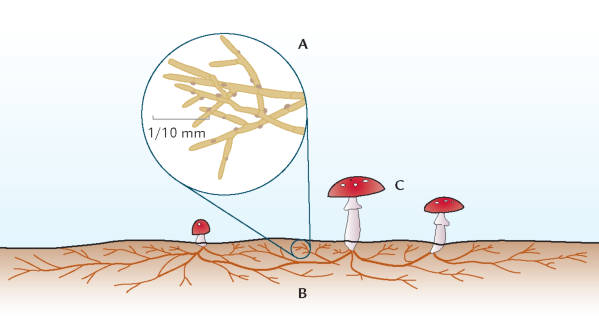
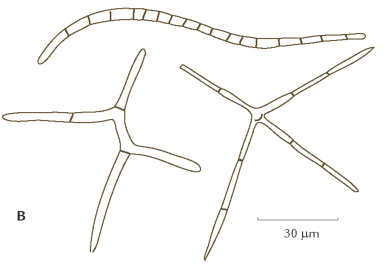
FIGUR 6-1. Mange svampe består basalt set af et netværk af tynde hyfetråde, der typisk er 10 μm (1/100 mm) tykke (A). Tilsammen udgør disse hyfer svampens mycelium (B), hvorfra der dannes de frugtlegemer (C), som man normalt betragter som svampe. Frugtlegemerne varetager en vigtig del af svampens livscyklus, nemlig dannelsen af kønnede spredningsenheder, de såkaldte sporer. De små øjer på tegningen af svampens hyfer er såkaldte øskner, der hos mange svampe findes, hvor to celler i en hyfe er adskilt af en cellevæg.


FIGUR 6-14. Svampesamfund tilknyttet muldbund er ofte særlig veludviklede langs vejkanter, hvor jævnlige forstyrrelser samt mineralholdigt vejgrus skaber et næringsrigt miljø, som bl.a. keglemørkhat sætter stor pris på.

FIGUR 6-15. Den meget smukke indigo-slørhat har en sydlig udbredelse i Europa og er meget sjælden i Danmark. Den synes her i landet at være tilknyttet sydvendte skrænter i skove på plastisk ler, hvor lejlighedsvise udskridninger skaber et ganske specielt, lysåbent miljø med blotlagt mineraljord.

FIGUR 6-16. Tragtformet læderpigsvamp er sammen med flere andre mykorrhizasvampe tilknyttet næringsfattig sandbund. Den er meget følsom over for kvælstofnedfald fra transport og landbrug og findes nu kun i de yderste kystegne, hvor nedfaldet af kvælstof fra atmosfæren er minimalt.
Skovens skjulte netværk
Det forhold, at svampe er opbygget som mycelier af mikroskopisk tynde hyfer, er af afgørende betydning for deres succes som nedbrydere, parasitter og symbionter. Svampehyfer kan uden problemer vokse ind i og gennemvæve jord, plantevæv og andet organisk materiale, hvor de ved hjælp af udskilte enzymer eller særlige sugeorganer (se senere) kan optage næringsstoffer fra deres omgivelser.
De mikroskopiske hyfer gør, at svampene ofte betragtes som mikroorganismer, men denne betragtning er ikke dækkende for forståelsen af svampenes betydning i skovøkosystemet. De vidstrakte mycelier gør svampene i stand til at transportere næringsstoffer vertikalt og horisontalt i skovbunden, f.eks. fra en falden bøgestamme og hen til rødderne af et levende træ. Svampene kan derfor betragtes som netværksdannende organismer, en slags skovens infrastruktur, der har en del til fælles med det netværk af kabler, gas- og vandledninger, der gennemvæver grunden under et typisk villakvarter.
Det skal dog nævnes, at visse svampe ikke har mycelier, men lever helt eller delvist som gær. Dvs., at de består af mikroskopiske, runde eller aflange enkeltceller, der deler sig ved knopskydning. De fleste gærsvampe lever af simple sukkerstoffer og er tilknyttet specielle levesteder, f.eks. gærende væsker, overfladen af blade, blomsternektar eller indre organer hos dyr og mennesker, hvor de hyppigt optræder som parasitter eller symbionter.
Svamperiget

FIGUR 6-2. Videnskaben beskriver med jævne mellemrum nye svampearter på baggrund af fund fra Danmark. I efteråret 2006 kunne svampeforskeren Tobias Frøslev således offentliggøre fund af fire nye knold-slørhatte-arter (tilhører slægten Cortinarius, underslægt Phlegmacium) fra Danmark. En af de nye arter er Cortinarius vesterholtii, opkaldt efter en anden dansk svampeforsker, Jan Vesterholt. Arten kendes indtil videre fra Møns Klinteskov og Marselisborg Skov syd for Århus.
Svampene er traditionelt blevet betragtet som en slags sære planter uden fotosyntese og med sporer som det vigtigste middel til formering og spredning. Svampene udgør dog deres helt egen, afgrænsede organismegruppe, der nu samles i et selvstændigt biologisk rige på linje med planteriget og dyreriget. Studier af svampes arvemateriale og cellestrukturer har vist, at svamperiget meget tidligt i evolutionen skiltes fra dyreriget, hvor de dyriske svampe, f.eks. ægte vaskesvamp, sjovt nok ser ud til at være de nærmeste slægtninge. Svampene er med andre ord nærmere beslægtet med dyr end med planter. Svampene er faktisk så afvigende fra både planter og dyr, at man må udvide de overordnede samlebetegnelser flora og fauna med en tredje enhed, der alene omfatter svampene. Denne enhed kaldes her i bogen funga efter det latinske ord for svamp, fungus.
Man kender ikke det nøjagtige antal svampearter i verden. Der er beskrevet ca. 100.000 arter, men man mener, at der er over en million. Alene fra Danmark kendes mindst 6.000 svampearter, men det reelle artsantal kan meget vel vise sig at være langt større, da mange svampegrupper er mangelfuldt undersøgt. Hvert år bliver adskillige arter erkendt for første gang i Danmark, og der beskrives jævnligt nye arter, også paddehatte med kødede frugtlegemer, på baggrund af svampefund i den danske natur (figur 6-2).
Skoven er den naturtype i Danmark, der uden sammenligning huser den mest artsrige funga. Man kan ikke finde en skov uden svampe, og skovens økologi kan ikke forstås, uden at svampene tages med i billedet: Svampene er nøglespillere ved planternes optagelse af vand og næringsstoffer i skovbunden; de vokser skjult i det grønne løv i trækronerne, og de er essentielle i skovens nedbrydningsprocesser. Også for mange dyr spiller samliv med svampe en afgørende rolle. De fleste vedboende insekter kan således ikke selv fordøje veddets hovedbestanddele, cellulose og lignin, men udnytter i stedet svampenes evner i den henseende. Svampenes rolle er dog ikke kun positiv for skovens øvrige organismer. Mange svampearter snylter på andre levende organismer, så disse svækkes eller dør.
Fødeoptagelse
Svampe har i modsætning til planter ikke fotosyntese og kan derfor ikke selv danne kulhydrater. Ligesom dyrene er de derfor afhængige af at kunne optage kulhydrater, som planterne har produceret, enten direkte fra levende eller døde planter eller indirekte via organismer højere oppe i fødekæden.
Svampe mangler i modsætning til de fleste flercellede dyr et tarmsystem. I stedet må de indtage vand, kulhydrater og andre næringsstoffer direkte fra deres omgivelser. Det kan de gøre på flere måder, og manglen på mund og mave er ikke ensbetydende med, at svampene har et passivt fordøjelsessystem.
Nedbrydere
Mange svampe lever af at nedbryde dødt organisk stof, herunder komplekse forbindelser som cellulose og lignin. Disse nedbrydersvampe udskiller reaktive enzymer til omgivelserne gennem deres cellemembraner. Derefter spalter enzymerne de komplekse organiske stoffer til simplere stoffer, hvis molekyler er så tilpas små, at de kan passere ind i svampens celler og dér indgå i svampens stofomsætning. Man kan med andre ord sige, at nedbrydersvampene fordøjer deres føde frit i det omgivende miljø.
Nedbrydersvampe optræder i alle skovmiljøer og omfatter mange forskellige typer af svampe – fra mug og skimmel, der især findes i jord og på sukkerholdige substrater, til bladhatte og poresvampe, der er velkendte nedbrydere på nedfaldent løv, dødt ved og andre døde plantedele. Nedbrydersvampene og deres store betydning for skovens økosystem omtales nærmere i kapitlerne Nedbrydning og Dynamik og forstyrrelser i skoven.
Mykorrhiza
Andre svampe lever sammen med levende planter. Deres hyfer inficerer rødderne hos planterne og danner det, man kalder mykorrhiza eller svamperod. Samlivet kommer i stand ved, at svampehyfer og planterødder vokser tæt sammen, således at vand og næringsstoffer kan udveksles. Planten forsyner svampen med kulhydrater, mens svampen kvitterer med næringsstoffer og vand. Normalt er mykorrhiza til gavn for både plante og svamp. Svampens mycelium kan forlænge plantens rodlængde betragteligt, og de meget tynde hyfer har den fordel, at de kan optage vand og næringsstoffer fra langt finere porer i jorden end de meget grovere planterødder er i stand til.
Afsnittet fortsætter efter boksen.
Boks

A (1). Bøgerod med forskellige ektomykorrhizaer. Bl.a. ses en sort, nubret type af frynsehinden Tomentella bryophila, en sølvhvid type med mycelietråde af gulmælket skørhat og en glat, gul type dannet af en foldhat.
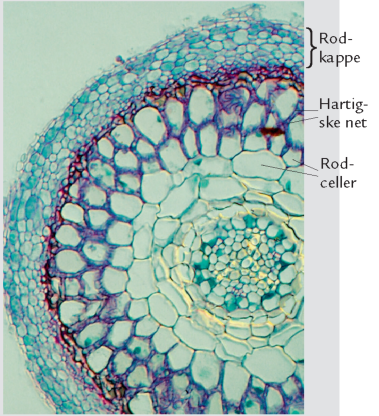
A (2). Tværsnit af bøgerod med ektomykorrhiza.
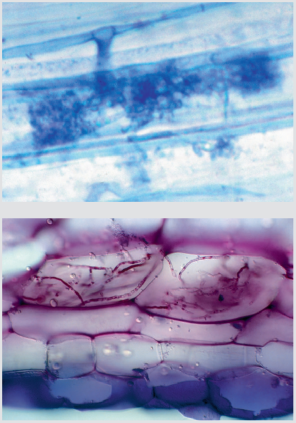
B. Øverst grenet arbuskel dannet af en Glomus-art i en plantes rodcelle. Farvet med trypanblåt. Nederst nøgler af svampehyfer i rodceller hos maj-gøgeurt. Farvet med toluidinblåt.

FIGUR 6-3 (a). Biotrofe parasitter omfatter en række svampearter, der lever af de kulhydrater, deres plantevært danner ved fotosyntese, uden at give noget positivt igen. En velkendt gruppe er meldug, her ege-meldug (Microsphaera alphitioides), der danner tynde, melede eller duggede belægninger på blade af især yngre egetræer (A). Også rustsvampe kan give en række forskellige synlige symptomer hos angrebne planter, gerne i form af orange til rustbrune pletter eller udvækster på blade eller stængler. Her ses arten Puccinia sessilis på et blad af ramsløg (B).
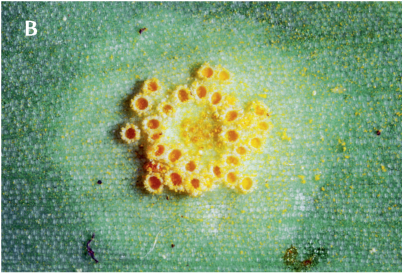
FIGUR 6-3 (b). Her ses arten Puccinia sessilis på et blad af ramsløg.

FIGUR 6-4. Grå snyltekølle, eksempel på en nekrotrof parasit, der angriber dyr (her en billelarve).
| Boks 6-1: Mykorrhiza Tv. bøgerod med forskellige ektomykorrhizaer. Bl.a. ses en sort, nubret type af frynsehinden Tomentella bryophila, en sølvhvid type med mycelietråde af gulmælket skørhat og en glat, gul type dannet af en foldhat. Th. Tværsnit af bøgerod med ektomykorrhiza. Foto: R. Kjøller. Mykorrhiza er et samliv mellem planter og svampe, der muliggør udveksling af næringsstoffer mellem de to parter. Ektomykorrhiza En række velkendte basidiesvampe, bl.a. kantareller, skørhatte og fluesvampe, men også sæksvampe, f.eks. trøfler, danner såkaldt ektomykorrhiza (figur A). Her lejrer svampehyferne sig tæt omkring de yderste cellelag i skovtræernes rødder. De danner en tæt kappe omkring de inficerede rodspidser og et netværk af hyfer, kaldet det hartigske net, mellem rodens ydre celler. Kun inderst mod rodmarven findes ingen svampehyfer. Mykorrhiza udgør koblingen mellem svamp og træ og er det knudepunkt, hvor svampen leverer næringsstoffer og vand til svampen, mens svampen til gengæld modtager kulhydrater fra træet. Rødder med ektomykorrhiza adskiller sig tydeligt fra ikke-inficerede rødder ved at være opsvulmede, rigtgrenede og korte. Ofte har de tillige en tydelig farve eller er forsynet med tråde og bundter af svampemycelie. De fleste af vore skovtræer, med ask og ahorn som væsentlige undtagelser, danner ektomykorrhiza, og i mange tilfælde er der tale om en værtsspecifik eller stærkt selektiv relation mellem svamp og træ. Den anden vej rundt er graden af specificitet lille, og alle ektomykorrhizadannende træarter har mange potentielle svampepartnere. Øverst grenet arbuskel dannet af en Glomus-art i en plantes rodcelle. Farvet med trypanblåt. Nederst nøgler af svampehyfer i rodceller hos maj-gøgeurt. Farvet med toluidinblåt. Foto: øverst J. Rosendahl, nederst H. Rasmussen. Arbuskulær mykorrhiza Hos ask og ahorn samt de fleste urter og buske findes arbuskulær mykorrhiza. Her vokser svampehyferne ikke kun imellem rodcellerne, men også ind i disse, hvor de danner grenede strukturer, de såkaldte arbuskler (figur B, øverst). Disse strukturer fungerer som koblinger mellem svamp og planter, således at vand og næringsstoffer kan udveksles. Til gengæld dannes der ingen kappe af svampehyfer rundt om rødderne, og man kan typisk ikke med det blotte øje se, om der er svampe i dem. Arbuskulær mykorrhiza dannes overvejende af mikrosvampe uden egentlige frugtlegemer, især kuglesvampe. Kuglevampene lever en meget skjult tilværelse som mykorrhizapartnere i jordbunden. Enkelte arter i gruppen kan danne trøffelagtige frugtlegemer, men generelt er der tale om en svampegruppe, der er usynlig for det blotte øje. Orkide-mykorrhiza Hos orkideer findes en særlig form for parasitisk mykorrhiza, som dannes ved, at hyfer af forskellige svampe vokser ind i det spirende orkide-frø. Her danner svampen tæt sammenvævede hyfenøgler i hver rodcelle (figur B, nederst). Disse hyfenøgler opløses gradvist, således at planten kan udnytte næringsstoffer fra svampen. Mykorrhizalivet er nødvendigt for, at de meget små orkidefrø kan få næring nok til at spire, men hos mange orkideer fortsætter samlivet i rødderne hele livet. Det gælder ikke mindst arter, der har meget lidt eller slet intet bladgrønt, f.eks. rederod og koralrod. Mange af de svampe, man har isoleret fra orkide-rødder, danner tillige ektomykorrhiza med skovtræer. Disse svampe lever således i to forskellige mykorrhizaforhold på én gang. Et mutualistisk, hvor et skovtræ forsyner svampen med kulhydrater til gengæld for vand og næringsstoffer, og et parasitisk, hvor svampen leverer kulhydrater videre til en orkide uden at få nogen igen. |
|---|
Afsnit fortsætter her.
Mykorrhiza dannes af en lang række forskellige svampe, både velkendte spisesvampe med kødede frugtlegemer og mikrosvampe, der lever et yderst skjult liv i skovbunden (boks 6-1).
Mykorrhizasvampenes mycelier deles ofte mellem flere træer, som derved er forbundet af et hyfenetværk. Man har påvist, at kulhydrater produceret af ét træ gennem dette netværk kan vandre over i et andet. Nogle forskere har ligefrem foreslået, at gamle træer med stor produktion af kulhydrater kan hjælpe skyggede artsfæller i konkurrencen mod andre træarter. Det er dog lige så sandsynligt, at de dominerende træer udnytter deres mindre heldigt stillede naboer.
Mange mykorrhizasvampe indgår direkte eller indirekte i skovbundens nedbryderfødekæder (se Nedbryderfødekæder og Svampe). Derved er skovens træer i direkte forbindelse med nedbryderfødekæden og kan med andre ord potentielt opsuge næringsstoffer direkte fra en tabt gren i skovbunden via deres mykorrhizasvampe. Netværket af indbyrdes interagerende mycelier af nedbrydende og mykorrhizadannende svampe betegnes ofte som et skovens “wood wide web” med klar reference til internettet.
Endofytter
I planternes grønne dele lever de såkaldte endofyttiske svampe. Endofyttiske svampe vokser i det levende plantevæv uden at give anledning til synlige symptomer. Der er tale om en uhyre artsrig, men meget dårlig kendt svampegruppe med flere forskellige økologiske strategier. Nogle endofytter vokser aktivt i deres levende vært, mens andre er til stede som passive hvilestadier. De aktiveres først, når planten eller plantedelen dør, hvorefter de fordøjer de lettilgængelige kulhydrater og proteiner, der er tilbage i de døde planteceller, eller ernærer sig som nedbrydere af cellulose, lignin og andre strukturstoffer. Den latente tilstedeværelse giver et stort forspring i forhold til nedbrydersvampe, der først inficerer plantematerialet, når dette er dødt (kapitlet Nedbrydning).
Parasitter
Der findes også svampe, som aktivt parasiterer blade, skud og forveddede plantedele, hvor de ofte giver meget synlige symptomer. Mange parasitiske svampe danner særlige sugeorganer, som vokser ind i levende planteceller, hvorfra de optager simple kulhydrater. Disse såkaldt biotrofe parasitter kan kun optage næring fra levende planter (figur 6-3). Andre svampe er nekrotrofe og dræber plantevæv over en bred front, for derefter at udnytte de frigjorte næringsstoffer. Disse nekrotrofe parasitter kan typisk leve i længere tid som rene nedbrydere på dræbte planter (Svampe og skovdød).
Mange symbiotiske og parasitiske svampe er snævert knyttet til en enkelt plante- eller dyreslægt eller sjældnere til en enkelt art og kaldes derfor værtsspecifikke (figur 6-4 og 6-5). Værtsspecificet udvikles, fordi svampen skal accepteres af plantens eller dyrets specifikke forsvarssystemer eller have midler til at overvinde disse.
Det kræver en høj grad af specialisering, der ofte er udviklet over millioner af år i et tæt samspil mellem svampen og dens vært.
Flere ernæringsformer samtidig
Der er ikke nødvendigvis skarpe skillelinjer mellem svampenes forskellige ernæringsmåder. For mange svampe er det evnerne som nedbrydere, der er forudsætning for deres samliv med dyr. Det gælder f.eks. svampe, der udnyttes af insekter til at fordøje dødt træ, og for mange mykorrhizasvampe, som med effektive enzymsystemer er i stand til at frigøre hårdt bundne næringsstoffer fra dødt, organisk stof i skovbunden.
Afsnittet fortsætter efter boksen.
Boks 2
|
FIGUR 6-5. Skæl-rørhat (Leccinum) er et eksempel på en svampeslægt med ektomykorrhiza og en høj grad af værtspecialisering. Rød birke-rørhat (A) findes således kun med birk, og orange aspe-rørhat (B) vokser udelukkende med bævreasp. Foto J. Vesterholt. |
|---|
Afsnit fortsætter her.
Spredning
Svampe kan sprede sig til nye voksesteder ved, at myceliet langsomt vokser sig større. Denne vegetative spredning er effektiv, men langsom og fungerer kun i nogenlunde homogene og stabile miljøer, f.eks. en uforstyrret skovbund, hvor svampen hele tiden kan finde noget at leve af. Næsten alle svampe danner tillige mikroskopiske spredningsceller, som sætter dem i stand til at sprede sig og overleve under ustabile og heterogene forhold. Disse spredningsceller kaldes sporer og kan dannes enten kønnet eller ukønnet.
Ukønnede sporer kan dannes ved simpel afsnøring af hyfeender eller fra specialiserede organer (figur 6-6), men er under alle omstændigheder genetisk identisk med deres ophav. Under de rette omstændigheder kan ukønnede sporer umiddelbart danne et fuldt funktionelt mycelium. Svampe med ukønnede sporer kan hermed så at sige klone sig selv, og genetisk identiske svampeindivider kan spredes over meget store områder. Bl.a. hos skimmelsvampe er en sådan klonal spredning med ukønnede sporer den helt dominerende spredningsform.
Afsnittet fortsætter efter boksen.
Boks 4
|
FIGUR 6-6. Spredning med ukønnede sporer ses hos mange forskellige svampe. Hos halvkugleformet kulbær dannes der store mængder af ukønnede sporer fra grå tottede, støvede strukturer (A), der siden gennemvokses af de rustrøde stroma, der indeholder kønnede frugtlegemer (B). Under mikroskopet ses de grenede hyfestrukturer, hvorfra de ellipseformede sporer frigives (C). Også hos barksvampen Botryobasidium aureum finder man ofte ukønnet og kønnet sporedannelse side om side. De kønnede sporer dannes i det tynde hvide spind (D), mens de gullige puder (E) er dannet af ukønnede citronformede sporer, der i massevis frigøres fra grenede hyfer (F). Foto A og B: T. Læssøe, D og E: Jacob Heilmann-Clausen, C: tegning fra Y-M. Ju & J.D Rogers. |
|---|
Afsnit fortsætter her.
Kønnede sporer dannes ved en celledelingsproces, hvorved der opstår nye kombinationer af arvematerialet. De kønnede sporer dannes oftest i komplekse, specialiserede strukturer, frugtlegemerne (se næste afsnit). Kønnede svampesporer indeholder ligesom menneskelige æg- og sædceller kun ét sæt kromosomer. Hos dyr og planter skelner man mellem hanlige og hunlige kønsceller og -organer, f.eks. sæd- og ægceller eller pollen og støvfang, men dette er meningsløst for de fleste svampes vedkommende. Svampe optræder ikke som hanner og hunner og har ikke støvfang, der skal befrugtes af frigivne sporer. Det betyder dog ikke, at svampe mangler et sexliv, tværtimod. I stedet for køn taler man om parringstyper hos svampene. Parringstypen bestemmes af et eller to gener, der hver især findes i mindst to varianter (alleler).
Afsnittet fortsætter efter boksen.
Boks 6
|
FIGUR 6-7. Svampesporer kan være særdeles forskelligt udformet med en størrelse, der typisk varierer fra 5 til 50 μm. De fleste sporer er tilpasset spredning med luftstrømme, og deres ringe størrelse betyder, at de kan svæve i luften og blive spredt over meget lange afstande med vinden (A). Dette forhold kan formodentlig forklare, at mange svampe har en meget vid, i visse tilfælde global, udbredelse. Der findes dog også svampe, hvis sporer først og fremmest spredes gennem vand (B). Det er ikke alle sporetyper, der er beregnet til spredning. Mange svampe danner således tykvæggede hvilesporer, der først og fremmest sikrer, at svampen kan overleve lokalt i en periode med ugunstige forhold, f.eks. stærkt udtørring eller mangel på levende værtsplanter i vinterhalvåret (C). Tegninger A: Fra Petersen, 1998, B: fra Lembke & Seerup, 1996, C: fra Langer, 1994. |
|---|
Afsnit fortsætter her.
Når en kønnet spore lander i et egnet miljø, vil den spire til en hyfe, der kan vokse og optage næring, men den kan kun danne nye, kønnede sporer, efter at den har forenet sig med et andet, uparret mycelium af samme art. Foreningen af to uparrede mycelier kan dog kun ske, hvis de bærer forskellige alleler af de gener, der bestemmer parringstypen. Hos mange svampe findes i dusinvis af forskellige alleler af disse gener, og de fleste møder mellem to uparrede mycelier kan derfor give ophav til en parring og dannelsen af et nyt mycelium, der igen kan danne kønnede sporer. Man kan med andre ord sige, at svampe ikke har to, men utallige køn.
Svampesporer er meget forskelligt udformede, men de er overvejende tilpasset en passiv spredning med luftstrømme eller vand (figur 6-7). Langt de fleste sporer går tabt uden at give ophav til nye mycelier. Det skyldes enten, at de havner på steder, hvor der ikke er betingelser for spiring og vækst, at de ædes af smådyr, eller at de dræbes på grund af længere tids udsættelse for UV-lys. Produktionen af sporer er derfor gigantisk hos mange svampe. Et enkelt frugtlegeme af tøndersvamp kan i løbet af et år producere flere billioner sporer, hvoraf kun en forsvindende lille andel vil spire til nye, levedygtige individer.
Nogle svampe har dog en mere målrettet og effektiv spredning over korte afstande. En del svampe spredes således med insekter eller pattedyr, der æder svampenes sporer, som er tilpasset til at kunne passere gennem dyrenes tarmsystem uden at miste spiringsevnen – det gælder bl.a. trøfler. Som belønning får dyrene næring fra sukkerholdige stoffer, der omgiver sporerne, eller fra de frugtlegemer, hvori sporerne findes (figur 6-8).
Afsnittet fortsætter efter boksen.
Boks 8
|
FIGUR 6-8. Almindelig stinksvamp (Phallus impudicus) danner sine sporer i en stærkt lugtende, kulhydratholdig og olivengrøn sporemasse, som tiltrækker mange forskellige insekter, ikke mindst fluer (A). Når fluerne æder af sporemassen, klæber nogle af sporerne til fluernes ben, hvorved de spredes til nye mulige voksesteder, når fluen flyver videre. En lignende spredningsstrategi anvendes i slægten Ophiostoma (B). Her udskilles både kønnede og ukønnede sporer i en slimet, kulhydratholdig væske, der ædes af eller klæber til forskellige insekter. Foto A: J. Heilmann-Clausen, B: J.H. Petersen. |
|---|
Afsnit fortsætter her.
Den relative betydning af vegetativ spredning og sporespredning er meget forskellig hos forskellige svampe. Nogle er helt afhængige af hyppig sporespredning. Det gælder specielt arter tilknyttet kortlivede, isolerede levesteder, f.eks. ekskrementer. Svampe tilknyttet meget stabile og vidt udbredte levesteder kan derimod blive meget store og gamle og er derfor mindre afhængige af sporespredning. I Canada har man fundet et mycelium af mørk honningsvamp, en art der også findes i Danmark, med en udstrækning på ikke mindre end 965 ha og en diameter på over 3 km. På baggrund af artens normale væksthastighed skønnes dette mycelium at have en alder på mellem 2.000 og 8.000 år. Det er således med nogen ret blevet fremført, at svampe hører til de største og ældste organismer på Jorden.
Det skal dog siges, at et så stort mycelium næppe fungerer som en biologisk enhed: Svampe har hverken et hjerte eller et centralnervesystem, der er nødvendige for individets enhed. Derfor kan svampemycelier uden videre splittes op i utallige adskilte enheder uden tab af funktionalitet, for siden hen igen at vokse sammen, hvis lejlighed bydes.
Opsplitning af svampemycelier i flere enheder og den udbredte forekomst af ukønnede sporer gør, at definitionen af et individ ikke er helt enkel hos svampene. Man har derfor indført begreberne ramét og genét for at holde styr på tingene. En genét er en gruppe af genetisk identiske individer dannet ved spredning af ukønnede sporer eller opsplitning af mycelier, mens en ramét betegner et enkelt individ inden for denne gruppe (figur 6-9).
Svampenes frugtlegemer
Hos mange svampe dannes de kønnede sporer i iøjnefaldende frugtlegemer, f.eks. de paddehatte man ser i skoven om efteråret. Man kan undre sig over, hvorfor svampene investerer i frugtlegemer, der i visse tilfælde kan veje adskillige kilo, når de ellers lever skjult i skovbunden og består af mikroskopisk tynde hyfer. Forklaringen er, at frugtlegemerne forbedrer svampenes mulighed for at få spredt deres sporer – men hvordan? En primær funktion af frugtlegemerne er at sikre, at sporerne kan nå op i turbulente luftlag over skovbunden, hvor luftstrømme kan føre dem væk fra udgangspunktet. Frugtlegemerne er desuden udviklet til at maksimere sporeproduktionen i forhold til den investerede biomasse. Der er mange måder at løse disse opgaver på, og svampefrugtlegemer kan være særdeles forskelligt udformet i overensstemmelse hermed.
Frugtlegemtyper
Afsnittet fortsætter efter boksen.
Boks 10
|
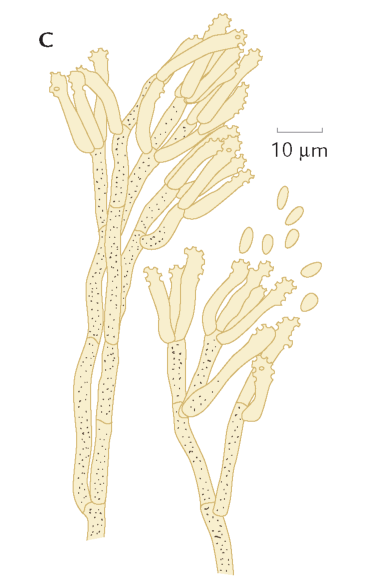
FIGUR 6-10. A) To sporesække, den højre med afskudte sporer. B) To basidier med sporer; på den højre er de netop afskudt ved den mekanisme, som vises i detaljer nedenfor (C). Efter Petersen, 1998. |
|---|
Afsnit fortsætter her.
Grundlæggende skelner man mellem to grupper af frugtlegeme-dannende svampe: sæksvampe (Ascomycota) og basidiesvampe (Basidiomycota) – opkaldt efter de celler, hvor sporerne dannes. Hos sæksvampene dannes de kønnede sporer i mikroskopiske sække, mens de hos basidiesvampe dannes på kølleformede celler, basidier (figur 6-10). Sæksvampenes sporesække fungerer som kanonrør i mikroverdenen. Ved modenhed frigøres sporerne eksplosivt, idet et væsketryk opbygget inde i sækken udløses. Kraften er tilstrækkelig til at slynge sporerne op til en halv meter op i luften, og under de rette forhold kan man både se og høre, hvordan hvidlige skyer af sporer hvislende frigøres fra sæksvampe gemt i skovbunden. Hos basidiesvampene sker sporefrigørelsen ved, at overfladespændingen i en vanddråbe ved sporens basis udløses. Denne mekanisme er betydelig mindre voldsom end sæk svampenes, men er dog tilstrækkelig til at bringe sporen nogle mikrometer væk fra basidien.
Både sporesække og basidier sidder tæt sammen i frugtlegemerne i et palisadeagtigt væv, der kaldes hymeniet. Dette hymenie er ofte meget forskelligt orienteret hos de to svampegrupper, netop på grund af forskelle i sporefrigørelsen (figur 6-11). Især hos basidiesvampene er hymeniet særdeles forskelligartet udformet for at øge mængden af basidier i forhold til frugtlegemets samlede størrelse (figur 6-12).
De fleste sæk- og basidiesvampe danner frugtlegemer, men der findes også svampe, som danner kønnede sporer uden egentlige frugtlegemer. De fleste rustsvampe danner således basidier fra helt simple strukturer, mens sækdug danner sporesække fra et udifferentieret væv på overfladen af angrebne blade og frugter. Derudover findes der tre store grupper af svampe, der helt mangler egentlige frugtlegemer, nemlig piskesvampe (Chytridomycota), koblingssvampe (Zygomycota) og Glomeromycota, som på dansk kunne kaldes kuglesvampe.
Afsnittet fortsætter efter boksen.
Boks 12
|
FIGUR 6-12. Rør, lameller og pigge er forskellige måder at opnå den forøgelse af overfladen på, som ses hos henholdsvis rørhatte (A), lamelsvampe (B) eller pigsvampe (C). Der findes yderligere et utal af mærkværdige frugtlegemetyper hos basidiesvampene, der bryder med disse overordnede mønstre, f.eks. støvbolde, koralsvampe og bævresvampe. Foto: J. Vesterholt. |
|---|
Afsnit fortsætter her.
Frugtlegemdannelse og årstider
De fleste forbinder svampe med efteråret, som da også er den årstid, hvor man har størst chance for at finde svampenes frugtlegemer i skoven. Men hvorfor er det sådan? Svampenes frugtlegemer repræsenterer en ganske betydelig investering og dannes derfor, når det bedst kan betale sig for svampen. To nøglefaktorer er vigtige her. Dels chancen for at frugtlegemet kan formå at sprede så mange sporer som muligt, dels chancen for at så mange af de spredte sporer som muligt kan spire til et nyt mycelium. Svampefrugtlegemer indeholder typisk 90 % vand, og et stabilt vandindhold er afgørende for, at sporerne kan frigives. Derfor finder man flest svampefrugtlegemer i fugtige perioder, typisk om efteråret, men også i højsommeren hvis bare nedbøren er stabil. Det er der ikke så mange, der er klar over, og mange habile svampesamlere går i våde somre glip af en stor høst af kantareller og karljohanner, fordi de ikke tager på svampejagt mellem badeturene ved stranden.
Langt færre svampe danner frugtlegemer om vinteren eller foråret. Disse arter må kæmpe med lave temperaturer, frost og især om foråret med tørke, der vanskeliggør et normalt stofskifte. Hos disse arter må det antages, at forhold omkring sporespiringen er årsagen til, at frugtlegemedannelsen sker uden for den gængse sæson. Der findes også en del svampe, hvor dannelsen af frugtlegemer og sporespredning ikke primært er bestemt af årstidernes vekslen. Især hos svampe tilpasset kortlivede levesteder, f.eks. gødning, bestemmes dannelsen af frugtlegemer f.eks. primært af nedbrydningsforløbet. Her sker dannelsen af frugtlegemer, når det enkelte svampeindivid har udnyttet sin føderessource fuldt ud, og inden eventuelle, mere konkurrencestærke arter tager over.
Storsvampe og mikrosvampe
Af praktiske årsager taler man ofte om storsvampe, der er kendetegnet ved at danne frugtlegemer af en vis størrelse (typisk større end 1 mm), og mikrosvampe, der mangler eller kun danner meget små frugtlegemer. Ifølge denne definition er piske-, koblings- og kuglesvampe mikrosvampe. Adskillelsen af de to grupper er betydelig mere problematisk inden for basidie- og sæksvampe, hvor arter med og uden frugtlegemer større end en millimeter ofte er nært beslægtede.
Opdelingen i storsvampe og mikrosvampe giver ikke megen mening i biologisk forstand, men kan være meningsfuld i praksis. F.eks. er det med enkelte undtagelser kun storsvampene, der behandles i populære svampebøger. Samtidigt er det værd at bemærke, at svampe med store frugtlegemer også har store og ofte vidtstrakte mycelier med en udbredelse, der skal måles i meter, mens mikrosvampe ofte, men ikke altid, har små mycelier, der ofte er mindre end en meter i udbredelse.
Skovens svampesamfund
Af de mere end 6.000 svampearter, der kendes i Danmark, vokser skønsmæssigt 3/4 eller ca. 4.500 arter overvejende i skove. Da mange svampegrupper, ikke mindst blandt mikrosvampene, endnu er mangelfuldt udforskede i Danmark, er den reelle artsrigdom i skovenes svampeverden dog langt større.
Adskillige forhold har betydning for fungaens sammensætning i forskellige skove, og man kan derfor ikke gå ud i en enkelt skov og finde samtlige arter. En vigtig nøglefaktor er skovens træartssammensætning. Det skyldes ikke mindst symbiotiske og parasitiske interaktioner mellem svampe og skovtræer, der medfører, at mange svampe kun kan leve sammen med bestemte værtstræer. Således findes kantareller, rørhatte, skørhatte og andre svampe, der danner ektomykorrhiza, ikke i skove af ask og ahorn. Disse træarter danner nemlig ikke ektomykorrhiza med sæk- og basidiesvampe, men har i stedet arbuskulær mykorrhiza med forskellige kuglesvampe. I mange tilfælde er koblingen mellem træværter og svampearter endnu mere snæver. Mange svampe med mykorrhiza eller en endofyttisk levevis er således tilknyttet ganske bestemte træarter og findes kun, hvor disse vokser (figur 6-5).
Også jordbundsforholdene har stor betydning for svampene. I skove på morbund med et veludviklet humuslag er svampebiomassen meget betydelig, men artsrigdommen er ikke nødvendigvis særlig stor, da det ofte er langlivede, konkurrencestærke arter, der findes på denne jordbund. Det er her, man skal gå på svampejagt, hvis man er ude efter karljohansvampe og brunstokket rørhat (figur 6-13), men også andre ektomykorrhizasvampe, ikke mindst skørhatte og fluesvampe er almindelige. Især sent på efteråret kan man desuden finde frugtlegemer af forskellige huesvampe, fladhatte og tragthatte, der nedbryder løv og nåle i skovbunden.
På muldbund er svampebiomassen typisk mindre, men ofte er artsrigdommen større. Det skyldes til dels, at den hurtigere stofomsætning og den større aktivitet af jordbundsdyr skaber et mere ustabilt og forstyrret miljø, der giver plads til, at en lang række ikke så konkurrencestærke arter kan etablere sig. Trævlhatte og foldhatte er blandt de ektomykorrhizadannere, der især findes på muldbund, ofte sammen med nedbrydere, f.eks. rødblade, parasolhatte og mørkhatte (figur 6-14).
En særlig artsrig og speciel funga findes i skove, hvor en kalkrig eller mineralrig, leret undergrund ligger tæt på overfladen. Især blandt de ektomykorrhizadannende svampe findes der en lang række arter, der kun findes sådanne steder, herunder mange koralsvampe, slørhatte og rørhatte med smukke og farvestrålende frugtlegemer (figur 6-15). Mange af disse arter har en sydlig udbredelse i Europa med nordgrænse gennem Danmark. Faktisk er det uklart, om det er rigdommen på kalk og andre mineralske næringsstoffer, der betinger forekomsten af disse arter, eller om det snarere er en kombination af lavt indhold af tilgængelige makronæringsstoffer (primært fosfor og kvælstof) og et særligt varmt mikroklima, der spiller ind. Ofte skal man nemlig ud på stejle, sydvendte skråninger, hvor solen bager sommeren igennem, og erosion blotlægger mineraljord med lavt indhold af fosfor og kvælstof, for at finde disse arter.
Også skove på sandbund uden et tykt humuslag, f.eks. i klitterræn, huser specielle svampesamfund tilpasset de ekstremt næringsfattige forhold (figur 6-16). Endelig findes der helt specielle svampesamfund i sumpskove, både i de skovbevoksede tørvemoser, hvor der er opbygget et tykt tørvelag, og i sumpskove med stående vand. Særlig ellesumpene huser en speciel funga, da el tilsyneladende ikke deler en eneste ektomykorrhizadanner med andre danske træarter. Svampesamfund tilknyttet bestemte skovtyper gennemgås nærmere i kapitlerne 14 til 18.
Kommentarer
Kommentarer til artiklen bliver synlige for alle. Undlad at skrive følsomme oplysninger, for eksempel sundhedsoplysninger. Fagansvarlig eller redaktør svarer, når de kan.
Du skal være logget ind for at kommentere.