
Omsætning af planktonalger
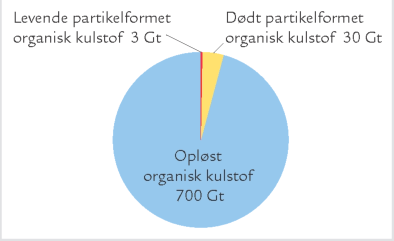
B. Mængden af de forskellige slags organiske stof i havet. Alle værdier er angivet i Gigaton (milliarder ton) kulstof.
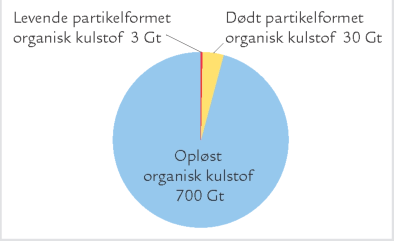
B. Mængden af de forskellige slags organiske stof i havet. Alle værdier er angivet i Gigaton (milliarder ton) kulstof.
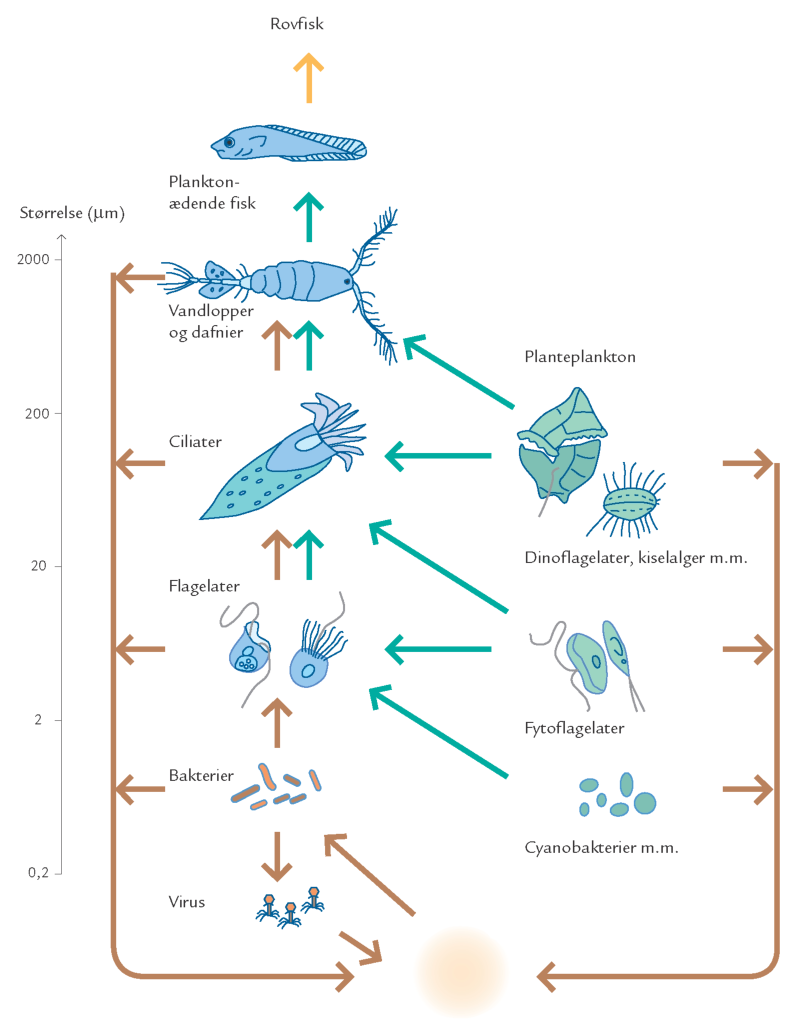
FIGUR 15-13. Skematisk model over fødekædens opbygning i de frie vandmasser. Organismerne er placeret efter størrelse med de mindste nederst og de største øverst. I højre kolonne er afbildet de autotrofe organismer (planktonalgerne) og i venstre side de heterotrofe organismer (bakterier, protozoer, dyreplankton og fisk). Pilene angiver transport af stof mellem organismegrupper, eller tab af stof til puljen af opløst organisk stof. Planteæderfødekæden er vist med grønne pile, mens nedbryderfødekæden er vist med brune pile. Den fødekæde, der baserer sig på dødt organisk stof, og som via bakterier danner grundlag for større organismer, kaldes den mikrobielle løkke.
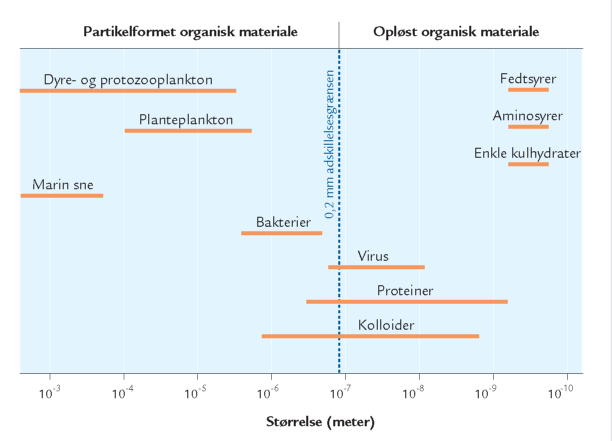
A. Størrelsesfordelingen af udvalgte organismegrupper og organiske stoffer i havets vandmasser. Grænsen 0,2 μm skiller mellem opløst stof og partikler. I denne fremstilling aftager størrelsen mod højre.
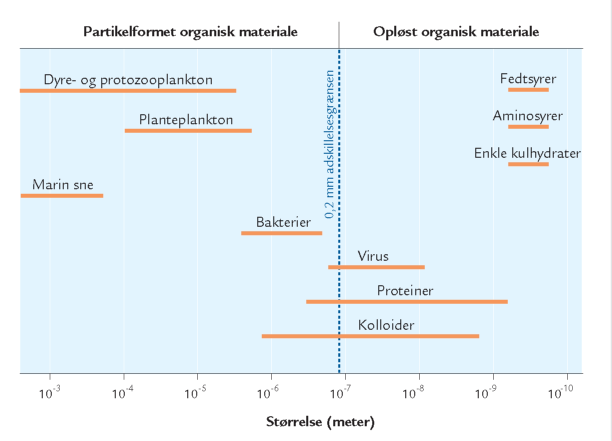
A. Størrelsesfordelingen af udvalgte organismegrupper og organiske stoffer i havets vandmasser. Grænsen 0,2 μm skiller mellem opløst stof og partikler. I denne fremstilling aftager størrelsen mod højre.
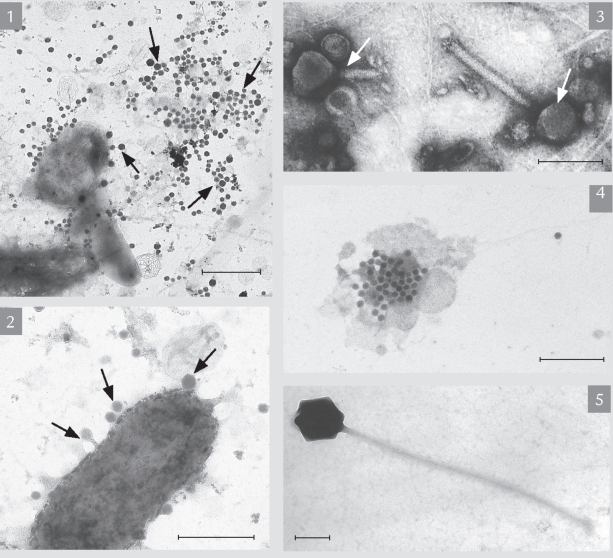
A. Virus indsamlet fra vandsøjlen og fotograferet gennem et elektronmikroskop. 1) Havvandsprøver med virus af mange forskellige former og størrelser (pile). 2) Bakteriecelle hvorpå forskellige viruspartikler har sat sig fast (pile) og er klar til infektion. 3) Viruspartikler under stor forstørrelse (pile). 4) Sprængt bakteriecelle, som de nye virus netop er ved at bryde ud af. 5) Ikke alle virus er lige små, her er en stor art. Målestok A, B & D: 1 μm. Målestok C & E: 0,1 μm.
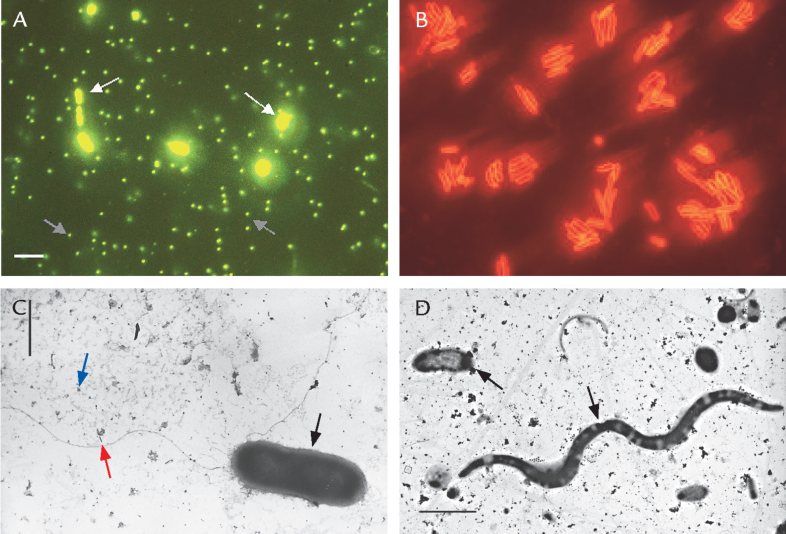
FIGUR 15-15. Bakterier fra havvand fotograferet med fluorescensmikroskop (A og B) og med elektronmikroskop (C og D). A) Bakterier i en havvandsprøve (hvide pile) farvet med et fluorescerende farvestof. De meget små prikker (grå pile) er virus. B) En kultur af havvandsbakterien Cellulophaga, hvis cellevæg er farvet med en antistof-farveteknik. C og D) Forskellige typer marine bakterier (sorte pile) fotograferet i elektronmikroskop. De små partikler (blå pil) er virus. Bemærk, at man kan se to flageller på den store bakterie i C (rød pil). Målestok 1 μm.
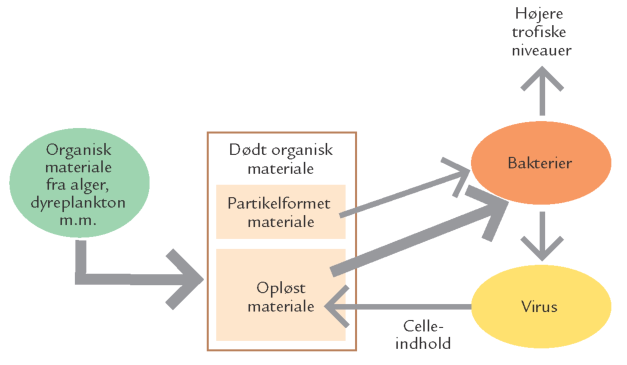
FIGUR 15-16. En skematisk fremstilling af bakteriers og virus' rolle i havets fødekæder.
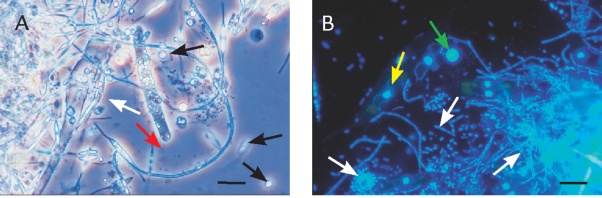
FIGUR 15-17. Nærbilleder af partikler af marin sne, der viser kolonisering med mikroorganismer. A) Foto fra lysmikroskopet af protozoer (sorte pile), der sværmer omkring en partikel bestående af detritus og algerester (bl.a. en død dinoflagellat, hvid pil). Bakterierne kan anes som små mørke streger (rød pil). B) Større forstørrelse af en partikel, som er farvet med det fluorescerende DNA-farvestof DAPI. Her ses tydeligt, at partiklen er overvokset af store mængder bakterier (hvide pile), der ligger enkeltvis, i kæder eller i klumper. Desuden ses bakterieædende, protozoer (grøn pil) og enkelte kiselalger med cellekerne og rød-brune kloroplaster (gule pile). Viruspartikler er for små til, at de kan ses ved denne forstørrelse. Målestok 10 μm.
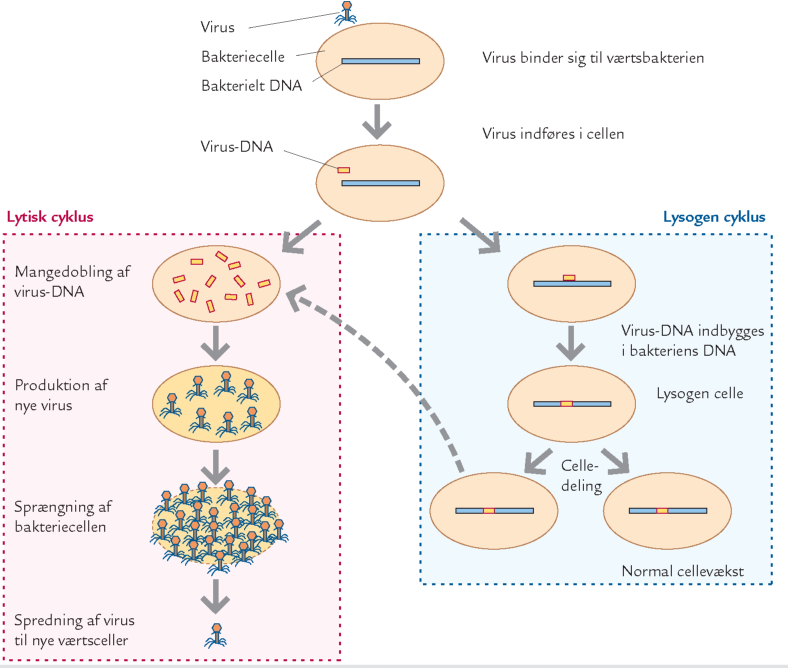
B. De to almindeligste livscykler for virus i havet: lytisk og lysogen cyklus.
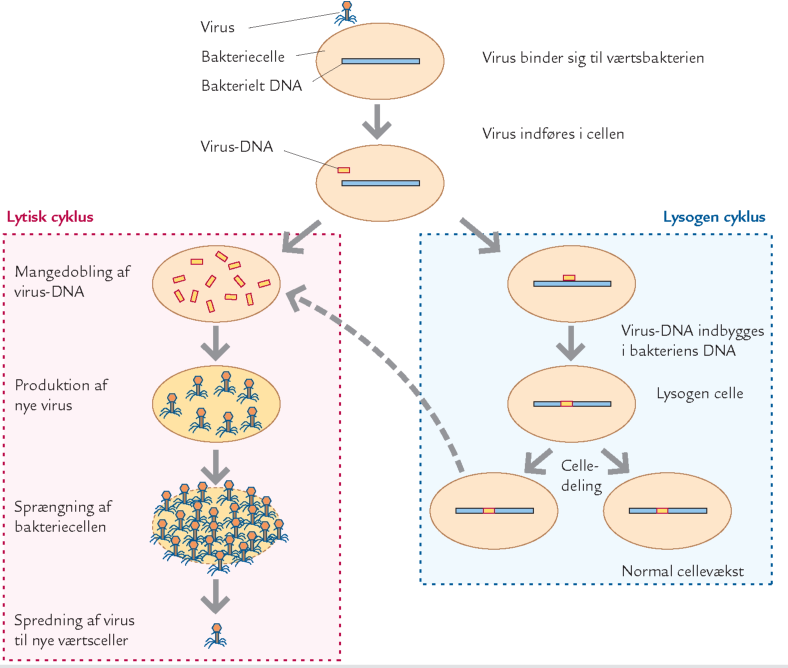
B. De to almindeligste livscykler for virus i havet: lytisk og lysogen cyklus.
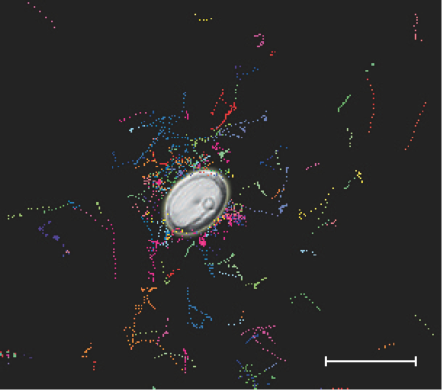
FIGUR 15-14. Illustration af hvordan bakterier sværmer omkring en død ciliat, der lækker organisk materiale og derved tiltrækker dem. De farvede, stiplede linjer er svømmespor fra enkelte bakterieceller. Hvert svømmespor afspejler en enkelt bakteries bevægelse i løbet af nogle sekunders videooptagelse med 0,04 sekunders mellemrum. Målestok 50 μM.
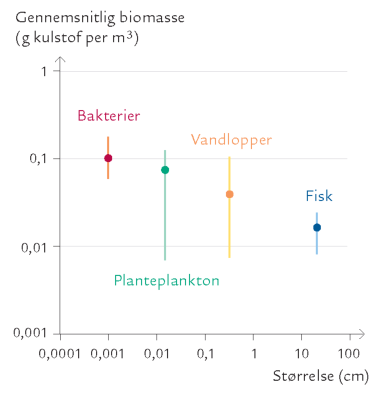
FIGUR 15-18. Størrelse af og biomasse for forskellige organismegrupper målt på en station i det sydlige Kattegat. Punkterne angiver den gennemsnitlige biomasse over et år. Værdien for fisk bygger dog på 25 års målinger. Stregerne angiver yderpunkterne for de enkelte målinger over perioden.
Primærproduktionens skæbne
Planktonalgernes produktion er fødegrundlaget for størstedelen af havets dyr og mikroorganismer. Det organiske stof, der er dannet ved fotosyntesen, og den energi, der er bundet i dette stof, omsættes i havets fødekæder, hvorved det udnyttes til vækst og formering af smådyr og fisk højere oppe i fødekæden. En del af vandsøjlens primærproduktion synker til bunds og indgår her i havbundens stofkredsløb (se Havbundens stofomsætning), men i reglen omsættes hovedparten af planktonalgernes produktion af organismer i selve vandsøjlen.
På grund af den stadige, turbulente vandbevægelse, der gør sig gældende i havet, vil små partikler støde sammen og danne større partikelansamlinger, såkaldte detritus-aggregater, der er synlige for det blotte øje. De kaldes for marin sne på grund af deres hvidlige, fnugagtige karakter (figur 15-9). Bakterierne i vandet menes at udskille de slimstoffer, der gør partiklerne klæbrige, så de forbliver sammen ved sammenstødene. Også intakte algeceller danner nu og da såkaldte fyto-aggregater, måske som følge af en egen slimudskillelse.
Afsnittet fortsætter efter boksen.
Boks
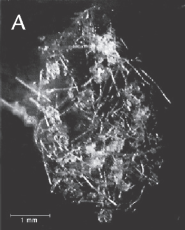
FIGUR 15-9 (a). Marin sne. Eksempler på millimeterstore sammenklumpninger af planktonisk materiale dannet i de frie vandmasser under påvirkning af vandets omrøring. Kiselalgeaggregater bestående af kæder af Chaetoceros sp. (A) og Skeletonema costatum (B), mens (C) består af dyreplanktonfækalier blandet med kæder af forskellige kiselalger.
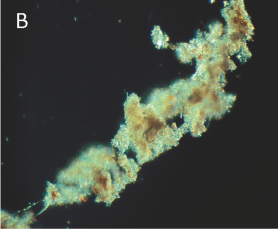
FIGUR 15-9 (b). Skeletonema costatum (B).
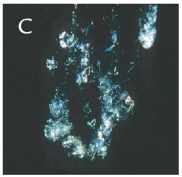
FIGUR 15-9 (c). Dyreplanktonfækalier blandet med kæder af forskellige kiselalger.
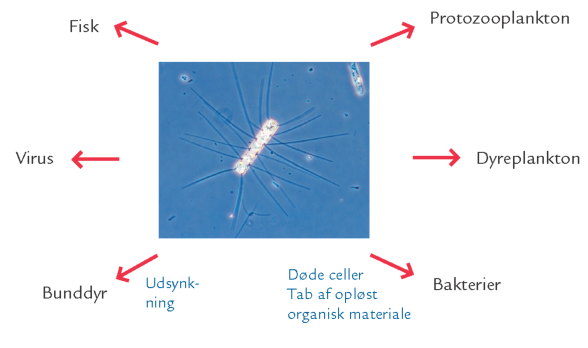
FIGUR 15-10. Planktonalgernes mulige skæbne.
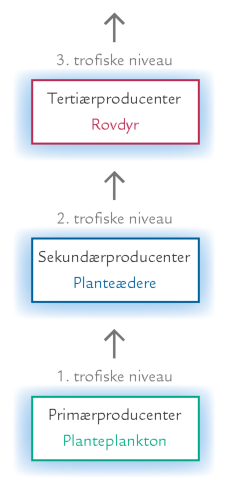
FIGUR 15-11. Trofiske niveauer i de frie vandmasser.
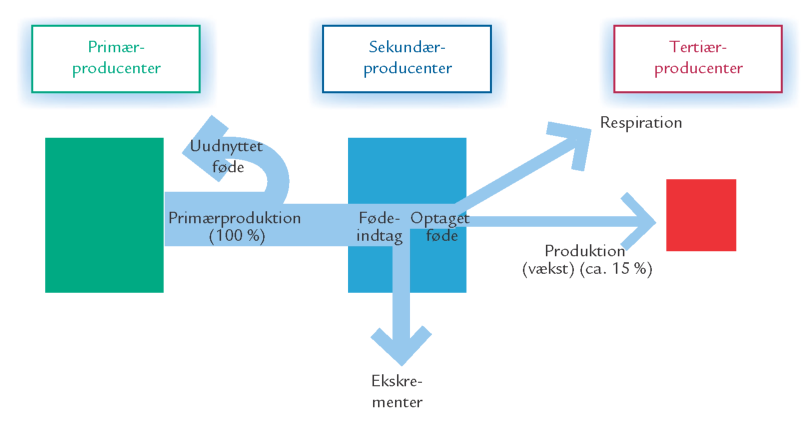
FIGUR 15-12. Transport og omsætning af stof igennem et trofisk niveau eksemplificeret ved planteædere.
| FIGUR 15-9. Marin sne. Eksempler på millimeterstore sammenklumpninger af planktonisk materiale dannet i de frie vandmasser under påvirkning af vandets omrøring. Kiselalgeaggregater bestående af kæder af Chaetoceros sp. (A) og Skeletonema costatum (B), mens (C) består af dyreplanktonfækalier blandet med kæder af forskellige kiselalger. Foto: T. Kiørboe, C. Lundsgaard og M. Olesen. |
Afsnit fortsætter her.
Da større partikler synker hurtigere end små, vil aggregerede partikler og især detritus-aggregater udgøre langt den største del af det materiale, der synker ned mod bunden. Af den samlede nedsynkning til havbunden er det derfor kun ca. 1/5, der synker ned som egentlige selvstændige planktonalgeceller, mens resten af nedsynkningen foregår som partikulært detritus.
I danske farvande vil detrituspartikler som følge af aggregatdannelsen gennemgående synke 10 gange hurtigere end frie planktonalger. På grund af aggregatdannelsen fjernes partikler og dermed næringsstoffer relativt hurtigt fra overfladevandet i de danske farvande, og vandet bliver ret klart i sommerhalvåret.
Hovedparten af planktonalgernes produktion omsættes imidlertid som nævnt i selve vandsøjlen. Her er der forskellige omsætningsveje: Cellerne kan enten blive ædt af dyreplankton eller fiskelarver, eller de kan dø som følge af næringsmangel eller virusangreb og efterfølgende blive nedbrudt af bakterier (figur 15-10). Derudover taber planktonalgerne opløst organisk materiale (boks 15-3), fordi både levende, aktivt voksende celler og døde celler er „utætte“. Opløst organisk materiale tabes også fra cellerne, når de går i stykker som følge af dyreplanktonets angreb eller som følge af virusinfektion. Sådanne tab svarer til omkring 5-25 % af primærproduktionen. Det opløste organiske materiale udgør en vigtig fødekilde for bakterier (figur 15-10 og 15-13).
Trofiske niveauer og omsætningens effektivitet i havet
Den biologiske produktion og omsætningen i havets frie vandmasser bliver ofte beskrevet som en sekvens af fødetrin (trofiske niveauer) fra planktonalger til de højere led i fødekæden (figur 15-11). I en sådan beskrivelse udgøres hvert trofisk niveau af organismer med samme funktion og angives med et nummer, der markerer afstanden til det første led i fødekæden, nemlig planktonalgerne.
Den lineære sekvens af trofiske niveauer, som vises i figur 15-11, er imidlertid en forenkling af de virkelige forhold i havet. De fleste organismer udnytter nemlig fødekilder fra flere forskellige trofiske niveauer og kan således ikke indpasses på et enkelt trofisk niveau. F.eks. kan nogle alger, bl.a. dinoflagellater, udføre fotosyntese, men kan også æde andre alger eller bakterier (kapitlet Det grønne plankton - planktonalgerne). Desuden agerer en stor del af dyreplanktonet som både rovdyr og planteædere, mens andre æder dødt materiale bestående af algerester og vandloppers ekskrementer. Endelig udnytter mange protozoer både alger og bakterier som føde, og nogle dyr skifter trofisk niveau i løbet af deres livscyklus. Til sidst kan det nævnes, at et trofisk niveau ikke behøver at omfatte levende organismer alene, der kan også indgå dødt organisk materiale.
Det samlede antal trofiske niveauer kan variere og afhænger bl.a. af primærproduktionens størrelse og af planktonsamfundets artssammensætning. På de øverste trofiske niveauer i vandsøjlen finder man voksne individer af rovdyr uden naturlige fjender. Ud over mennesker er det hajer, havpattedyr og visse blæksprutter.
På trods af den nævnte forenklings begrænsninger giver den en mulighed for at sætte tal på stof- og energistrømmene i havet. Ved at reducere de komplekse samspil imellem organismerne i havet til delvist forenklede sammenhænge, bliver det muligt at vurdere energistrømmene i forskellige havøkosystemer og sammenligne med forholdene i søer og på land.
Ved enhver fødeudnyttelse på et givent trofisk niveau sker der et tab af stof og energi i forbindelse med omsætning af føden (figur 15-12). Af den indtagne føde går kun en del til vækst. En stor andel (40-60 %) bruges i dyrenes energistofskifte (respiration) – dvs. den nedbrydes til kuldioxid og vand, og samtidig mistes en del af den bundne energi som varme. En mindre men varierende andel fordøjes ikke, men udskilles som ekskrementer. Omsætning af føden medfører desuden udskillelse af mineralske affaldsprodukter.
Normalt bliver mellem 10 og 50 % af den produktion, der findes på ét trofisk niveau, til ny produktion på næste niveau. Man kalder det overførselseffektiviteten. I eksemplet i figur 15-12 omdannes omkring 15 % af primærproduktionen til ny produktion af planteædere, der igen kan udnyttes af de mindre rovdyr. Selv om dette umiddelbart kan forekomme at være at stort tab, er det faktisk kendetegnende for overførslerne mellem de trofiske niveauer i vandsøjlen, at deres effektivitet er relativt høj.
Den høje effektivitet på 15 % i eksemplet skyldes bl.a., at planktonalgerne består af langt lettere nedbrydeligt materiale end landjordens primærproducenter. Vandets egenskaber og planktonalgernes ringe størrelse bevirker, at planktonalgerne kan holde sig „svævende“ og ikke i samme omfang som landplanterne behøver hårde støttevæv, som skal holde bladene op i lyset. Mens kun en relativt lille del af landplanternes primærproduktion reelt er tilgængelig for planteædere, kan langt hovedparten af planktonalgeproduktionen i vandsøjlen udnyttes.
Alt i alt betyder den højere effektivitet i omsætningen af primærproducenter i havet end på landjorden, at en relativ større andel af primærproduktionen i havet transporteres til de højere trofiske niveauer.
Afsnittet fortsætter efter boksen.
Boks 2
Dødt organisk materiale i havet forekommer i et bredt spektrum af størrelser fra enkelte, små molekyler (f.eks. glukose) til cm-store partikler som „marin sne“. Man skelner ofte mellem opløst organisk materiale og organisk materiale på partikelform; det partikelformede bliver tilbageholdt i et filter med porestørrelsen 0,2 μm, det opløste gør ikke (figur A). Opdelingen er praktisk, men siger ikke noget om stoffernes kemiske sammensætning, oprindelse eller økologiske rolle. Hovedparten af havets organiske materiale er opløst, faktisk over 95 % (figur B). Koncentrationen af opløst organisk kulstof i havet varierer fra under 1 mg C per liter i næringsfattige oceaner til 6-7 mg C per liter i næringsrige kystområder. Til sammenligning er der i en liter havvand typisk omkring 25 mg C bundet som uorganisk kulstof i kuldioxid, bikarbonat og karbonat, mens biomassen af planktonalger sjældent overstiger 1 mg C, selv ikke i meget næringsrige områder. Opløst organisk kulstof udgør således en enorm pulje af organisk stof på kloden, og mængden svarer omtrent til landjordens samlede plantebiomasse. Opløst organisk materiale omfatter både lavmolekylære stoffer som aminosyrer og sukkerstoffer og højmolekylære forbindelser som proteiner og langkædede sukkerstoffer, samt virus og andre meget små partikler på kolloidform. Det opløste organiske materiale synker ikke til bunds, men forbliver i vandsøjlen, hvor det udgør den vigtigste ernæringskilde for bakterierne.
|
|---|
|
B. Mængden af de forskellige slags organiske stof i havet. Alle værdier er angivet i Gigaton (milliarder ton) kulstof. |
|
A. Størrelsesfordelingen af udvalgte organismegrupper og organiske stoffer i havets vandmasser. Grænsen 0,2 μm skiller mellem opløst stof og partikler. I denne fremstilling aftager størrelsen mod højre. |
Afsnit fortsætter her.
Havets fødekæder
Den forenklede beskrivelse af vandsøjlens organismer i trofiske niveauer kan, som gjort ovenfor, benyttes til at beskrive de overordnede energistrømme i havet. En mere korrekt beskrivelse af havets fødekæder kræver en mere detaljeret gengivelse af samspillet mellem organismegrupperne. Til dette anvendes ofte en model, hvor primærproduktionen helt overordnet kan omsættes ad to veje: enten via en planteæderfødekæde eller via en mikrobiel nedbryderfødekæde, den såkaldte „mikrobielle løkke“ (figur 15-13).
Planteæderfødekæden
I planteæderfødekæden ædes planktonalgerne direkte af dyreplankton. Det planteædende dyreplankton består af både encellede og flercellede former. Flagellater (2-20 μm), ciliater og dinoflagellater (20-200 μm) er de dominerende grupper i det encellede heterotrofe plankton, der primært ernærer sig af de mindste planktonalger og af bakterier. Det flercellede dyre plankton (0,2-2 mm) består af forskellige grupper af krebsdyr (bl.a. dafnier og vandlopper), hjuldyr og larver af bundlevende dyr. De lever fortrinsvis af de større planktonalgearter, men nogle arter er rovdyr, der æder andre planktondyr (se Det farveløse plankton - protozoer og dyreplankton).
Nedbryderfødekæden
I nedbryderfødekæden foregår omsætningen via bakteriel nedbrydning af dødt organisk materiale (boks 15-3). Bakterier er prokaryote organismer, dvs. de er uden cellekerne. De er typisk fra 0,5 til 2 μm store og forekommer i et enormt antal på ca. 1 milliard bakterieceller per liter havvand.
Bakterierne i havet er ofte bevægelige og kan med deres flagel opnå svømmehastigheder på 20-100 μm per sekund. Da de også kan orientere sig i næringsstofgradienter, kan de svømme mod områder eller partikler med en forhøjet næringskoncentration, f.eks. omkring en døende celle (figur 15-14).
Havets bakterier er som de eneste organismer i vandsøjlen i stand til at udnytte de opløste organiske stoffer som deres vigtigste næringskilde (figur 15-15). Det opløste organiske materiale i havet udgør derfor grundlaget for den mikrobielle nedbryderfødekæde (figur 15-16).
Det opløste organiske stof har en yderst blandet sammensætning (boks 15-3). De lavmolekylære stoffer kan optages direkte over bakteriens overflade, mens de højmolekylære stoffer må undergå en delvis nedbrydning uden for cellen af specielle enzymer i bakteriernes cellemembran, før de kan optages.
Afsnittet fortsætter efter boksen.
Boks 6
|
Virus er de mindste og mest talrige organismer, man kender. De er typisk mindre end 0,1 μm og består grundlæggende af et stykke genetisk information (DNA eller RNA) indesluttet i et proteinhylster, der beskytter det genetiske materiale og fungerer som fasthæftningsorgan ved infektion. Virus forekommer i meget stort antal i det marine plankton, typisk 10-30 gange så mange som bakterier, altså 10-100 milliarder viruspartikler per liter havvand. De tilbringer en del af deres livscyklus som parasitter inde i en værtscelle som f.eks. en bakterie- eller en algecelle. Virus er en helt enestående livsform, der adskiller sig fra alle andre organismer ved ikke at have noget eget stofskifte. Virus er derfor i sig selv biologisk uvirksomme partikler, der er afhængige af en værtscelles stofskifte for at kunne formere sig. De fleste virus i havet inficerer bakterier, de er såkaldte bakteriofager, men også eukaryote alger og protozoer inficeres. Når en virus møder en værtsbakterie, fastgøres den til specielle proteiner i værtscellens membran, de fungerer som receptorer for virus (se figur B). Herefter indfører virus sit genetiske materiale (DNA eller RNA) i bakterien, i hvis DNA det indbygges. Det indførte genetiske materiale „kortslutter“ bakteriens eget stofskifte og programmerer det til at producere nye virus inde i cellen. Efterfølgende nedbrydes værtscellens ydre membran (cellen „lyserer“ eller sprænger), og de nye virus, fra ca. 10 til mere end 1000 virus per inficeret celle, frigives til omgivelserne og kan inficere nye værtsceller. Sammen med de nye virus frigives desuden den døde celles indhold af organisk stof, der udgør en vigtig fødekilde for bakterier i vandet. Denne cyklus, fra infektion til frigivelse af nye virus, gennemløbes normalt over en periode, der svarer til bakteriens generationstid, dvs. fra timer til dage. I visse tilfælde sker produktionen af virus ikke umiddelbart efter infektionen af værtscellen, men virus-DNA'et kan ligge i værtens genetiske materiale uden at være aktivt og blive spredt til den inficerede celles døtreceller. Senere kan det blive aktivt som følge af en ydre påvirkning, f.eks. ændringer i lys, temperatur eller næring (en såkaldt lysogen infektion, figur B). Virus spiller en vigtig rolle i den mikrobielle nedbryderfødekæde. Op imod 30 % af bakterieproduktionen tabes nemlig dagligt som følge af virusangreb, hvilket er lige så meget som protozoerne æder. Derudover påvirker virus indirekte den mikrobielle omsætning af organisk stof. En stor del af bakterieproduktionen frigives nemlig igen som opløst organisk materiale som følge af virusangreb, og dette materiale omsættes igen af andre bakterier. Virussenes aktivitet bevirker altså, at en del af det organiske materiale genbruges flere gange inden for bakte riesamfundet. Det øger energiforbruget i nedbryderfødekædens nederste led og gør, at der bliver mindre energi at overføre til større dyr i toppen af fødekæden. En anden vigtig egenskab ved virus er, at de er specifikke i deres valg af værtscelle, så en bestemt type virus eksempelvis kun kan inficere en bestemt art af bakterier. Virusinfektion kan reducere antallet af en enkelt alge- eller bakterieart i løbet af få timer, og virus kan derfor have stor indflydelse på overlevelsen af individuelle bakterie- og algearter. Virus kan derfor forme artssammensætningen i de mikrobielle samfund og påvirke den mikrobielle omsætning og diversitet i havet.
|
|---|
|
B. De to almindeligste livscykler for virus i havet: lytisk og lysogen cyklus. Efter Riemann & Middelboe, 2000. |
Afsnit fortsætter her.
Trods deres store antal, er bakteriernes biomasse i havet oftest kun nogle få μg C per liter. Bakteriebiomassen er altså 500-1000 gange mindre end koncentrationen af opløst organisk kulstof, der som nævnt i boks 15-3 ligger på fra under 1 til omkring 6-7 mg C per liter. En vigtig grund til, at bakteriebiomassen er så relativt lille, er, at kun 10-20 % af det opløste organiske materiale er umiddelbart tilgængeligt for bakterierne. Resten er unedbrydeligt eller meget svært nedbrydeligt, og det kan ikke omsættes biologisk inden for en kortere tidshorisont, men nedbrydes langsomt af især Solens uv-stråling.
Dødt organisk stof i partikelform udgøres bl.a. af „marin sne“ og vandloppeekskrementer. De døde partikler koloniseres af bakterier og nedbrydes langsomt på deres vej mod bunden. I forbindelse med denne bakterielle kolonisering af organiske partikler kan der dannes små mikrosamfund af bakterier, protozoer og virus på partiklerne (figur 15-17).
Bakterierne udgør i sig selv en vigtig fødekilde for flagellater og ciliater. På den måde bliver en del af det opløste stof via bakterierne kanaliseret tilbage til de højere led i fødekæden (figur 15-13). På grund af de mange led i den mikrobielle fødekæde sker der et stort tab af stof og energi, og en relativt lille del af det producerede opløste organiske stof ender som føde for det flercellede dyreplankton. I perioder med få og små planktonalgeceller kan protozoer dog udgøre en forholdsvis stor andel af føden for det større dyreplankton. I sådanne situationer bliver den mikrobielle fødekæde et vigtigt bindeled mellem planktonalgerne og det flercellede dyreplankton.
Planktonorganismernes betydelige tab af organisk materiale til vandet betyder, at 10-50 % – i nogle tilfælde endnu mere – af primærproduktionen omsættes af bakterier i vandet. Uanset at bidraget fra den mikrobielle nedbryderfødekæde til dyreplanktonets fødegrundlag varierer, så er den mikrobielle nedbryderfødekæde til enhver tid af stor betydning for den samlede omsætning i vandsøjlen.
Størrelsens betydning
Økologiske systemer er komplicerede – de omfatter hundredvis eller tusindvis af arter, der påvirker hinandens bestande og omgivelsernes kemi på forskellige måder. Artsrigdommen og variationen i tilpasninger er for planktonorganismernes vedkommende beskrevet i kapitlerne 4 og 5. Trods den store kompleksitet kan man alligevel finde en slags orden og regelmæssighed, der gælder for f.eks. alle planktonsamfund. Man kan endog opnå en forståelse af sådanne mønstre på grundlag af nogle få og enkle principper, f.eks. angående de egenskaber, der knytter sig til organismers størrelse.
Størrelsen af de enkelte organismer stiger mange størrelsesordener fra bakterier over encellede alger og protozoer, de mm-store, flercellede dyreplanktonorganismer til fisk, blæksprutter og havpattedyr. En hval er ca. 10 millioner (107) gange længere – og vejer dermed ca 1021 gange mere – end en bakterie. To forhold er her afgørende.
Det ene er, at organismers energistofskifte ikke øges proportionalt med deres størrelse, men snarere proportionalt med W3/4‘, hvor W er kropsvægten. Det vil f.eks. sige, at hvis man sammenligner to organismer, hvor den ene har en kropsvægt, der er 10.000 gange større end den anden, så vil den store organisme kun have et stofskifte (målt f.eks. som iltoptagelse) der er 1000 gange større end den lille organisme. Ydermere er væksthastigheden og dermed formeringshastigheden tæt koblet til energistofskiftet. Små organismers potentielle bestandstilvækst er langt højere end den, der gælder for store organismer. Dette indses ved at betragte generationstider. For bakterier er den omkring en time, for encellede eukaryoter mellem nogle timer og et døgn, for dyreplankton varierer den mellem et par døgn og nogle uger, mens fisk har en generationstid fra et til flere år.
Det andet forhold er, at i fødekæder er det større organismer, der æder mindre organismer. Som en lidt grov generalisering er rovdyr 10 gange større end deres byttedyr – her er mange undtagelser, men som en gennemsnitlig størrelse er det omtrent rigtigt. Som tidligere omtalt tabes der energi ved hvert led i fødekæden. Igen som en lidt flot generalisering kan man sige, at når et rovdyr æder 100 g byttedyr, vil det selv vokse med 15 g.
Med disse betragtninger kan man vise, at den totale biomasse af organismer skulle være omtrent den samme for alle størrelsesklasser. Dvs. at i en km3 havvand skulle den totale vægt af f.eks. planktonalger (størrelsesklassen 2-20 μm) være omtrent den samme som den totale vægt af hvaler og store fisk (størrelsesklassen 2-20 m).
Figur 15-18 viser et gennemsnitligt størrelsesspektrum for nogle størrelsesgrupper af organismer i Kattegat. Det ses, at hver logaritmisk størrelsesklasse har en biomasse på et sted mellem 0,1 og 0,01 g per kubikmeter vand – dog sådan, at de mindste organismer har en lidt højere biomasse end de største.
På grund af sæsonvariationen, der jo drives af vores tempererede klima, varierer størrelsesspektret noget gennem året. Der er altså krusninger på det generelle størrelsesspektrum, der er vist på figur 15-18. Ikke desto mindre viser eksemplet, at man kan finde enkle og forståelige mønstre i en tilsyneladende meget kompliceret natur.
Vejviser
Værket Naturen i Danmark i fem bind udkom i årene 2006-2013. Teksten ovenfor er kapitlet Omsætning af planktonalger.
- Forrige afsnit er Vandmassernes primærproduktion
- Næste afsnit er Havbundens stofomsætning
Kommentarer
Kommentarer til artiklen bliver synlige for alle. Undlad at skrive følsomme oplysninger, for eksempel sundhedsoplysninger. Fagansvarlig eller redaktør svarer, når de kan.
Du skal være logget ind for at kommentere.